Прическа «Колосок» своими руками фото
Прическа «колосок» была и остается быть любимой прической уже на протяжении долгих лет среди девочек и девушек. Конечно, она не всегда была такой популярной прической и уступала по моде другим. Но, всё модные тенденции, которые со временем уходили в тень, как известно, возвращаются и под влиянием новых порядков моды слегка видоизменяются. Прическа «колосок» снова популярна благодаря простоте её исполнения.
Такое же возвращение и обновление и произошло со всеми известной прической колосок». Если раньше колосок плели идеально гладко, то сейчас мы можем увидеть его в совершенно разных вариациях, иногда с эффектом и небрежности, будто девушка только что вскочила с кровати и на скорую руку заплела колосок, чтобы не опоздать в университет. Эта небрежность в такой изящной прическе нечто новое, модное и увлекательное. Сейчас эта прическа «дышит» свободой, которая придает ей мягкость.
Раньше прическа «колосок» считалась женственной и изящной прической с деловым оттенком, то есть другими словами, эта прическа соответствовала деловому стилю.
Сейчас же это современная прическа «колосок» или французская коса носит демократичный характер, так как она сочетается с различными стилями. Благодаря вариативности данной прически вы можете её сделать для дружеской встречи, романтической прогулки, деловой встречи, выхода в свет.
Прическа колосок своими руками фото:
Прическа «колосок» поэтапно
Прическа «колосок» поэтапно
Прическа «колосок»
Прическа «колосок» поэтапно
Прическа «колосок»
Молодые особы могут спокойно выбирать, в какой вариации они исполнят прическу «колосок». Например, они могут заплести два колоска сразу, то придаст их образу нечто новое, игривое. Или же романтичным девушкам как нельзя, кстати, подойдет такая прическа для свидания с возлюбленным. Один их вариантов романтичного колоска: необходимо взять спереди небольшую прядь и заплести колосок, заводя его за ухо. После того как вы колосок готов мы его место крепления закрываем свободными волосами. Чтобы придать прическе романтические нотки, волосы завиваются любыми способами, которые больше всего нравятся вам.
«Рыбий хвост» — это ещё один из вариантов плетения колоска. Рыбий хвост выглядит весьма эффектно и подходит для любого случая. Довольно часто такое плетение можно увидеть на невестах. Благодаря его небрежности и слегка растрепанности создается романтичный и невинный образ девушки. Большинство девушек отмечают, что эта прическа довольно долго держится на волосах, что явно является её плюсом.
Техника плетения прически «колосок».
В плетении прически «колосок» нет ничего ложного. Любая мама сможет с легкостью этому обучить свою дочку, чтобы та сама себе могла её заплетать.
Рассмотрим самое обыкновенноё плетение полоска. Для начала необходимо расчесать волосы. Затем берется прядь волос у лба, и делиться на три части. После этого начинаем плести косичку. Не забываем, что в этом плетении нам надо к пряди, которая оказывается с боку, вплетать дополнительную прядь волос с этой же стороны. Считается, что чем чаще делать такие захваты дополнительных прядей, то тем интересней получиться колосок. Когда колосок готов, то оставшиеся волосы можно либо заплести в хвост, либо доплести их уже обычной косичкой.
Когда колосок готов, то оставшиеся волосы можно либо заплести в хвост, либо доплести их уже обычной косичкой.
Как разнообразить плетение колоска
Во время плетения колоска не обязательно придерживаться идеального его выполнения, это даже вовсе делать не надо. Также чтобы не быть скучными и заурядными придумайте свой вариант плетения прически «колосок» или же воспользуйтесь нашими советами.
• Вы можете заплести вместо оного колоска сразу несколько.
• Можно заплести колосок в виде ободка вокруг головы.
• Не надо при добавлении в колосок брать все волосы, можно оставить по бокам несколько тонких аккуратных прядей, которые будут обрамлять лицо. Можно подкрутить пряди при помощи плойки.
• Заплети колосок на изнанку. Такое плетение будет выглядеть очень оригинально и интересно.
• Ниспадающий колосок. Такой колосок плетется очень просто. Не надо убирать все волосы, достаточно взять с любой стороны небольшую прядь и плести её сверху вниз, дополнительные волосы будут добавляться только сверху. Таким образом, по окончанию плетения у вас получиться что-то наподобие кармашка, если провести рукой под колоском. Остальные волосы должны быть гладкими и прямыми.
Таким образом, по окончанию плетения у вас получиться что-то наподобие кармашка, если провести рукой под колоском. Остальные волосы должны быть гладкими и прямыми.
• Можно заплести по тоненькому колоску с двух сторон на распушенных волосах. Не надо доплетать эти колоски до самого конца. Когда вы дойдете до затылка, то их надо скрепить вместе. Там где они будут соединены, можно закрепить в качестве украшения цветок и другие подходящие заколок.
• Еще можно с двух сторон заплести по колоску, которые на месте своего соединения будут заплетаться в узел или пучок. Так же можно колоски закончить косичкой.
• Заплетите колосок по низу, вплетая в него красивую ленту.
Самое главное при плетении колоска проявлять свою фантазию. И если не получилось создать что-то необыкновенное с первого раза, то не стоит отчаиваться, а надо пробовать ещё раз.
Задержка развития прикорневых колосков у пшеницы объясняет их частую осыпаемость соцветий и рудиментарную природу
%PDF-1. 7
%
1 0 объект
>/Метаданные 4 0 R/Страницы 2 0 R/StructTreeRoot 3 0 R/Тип/Каталог/ViewerPreferences 5 0 R>>
эндообъект
4 0 объект
>поток
Приложение Microsoft® Word для Microsoft 365/pdf
7
%
1 0 объект
>/Метаданные 4 0 R/Страницы 2 0 R/StructTreeRoot 3 0 R/Тип/Каталог/ViewerPreferences 5 0 R>>
эндообъект
4 0 объект
>поток
Приложение Microsoft® Word для Microsoft 365/pdf
0000000
конечный поток
эндообъект
2 0 объект
>
эндообъект
3 0 объект
>
эндообъект
5 0 объект
>
эндообъект
121 0 объект
>
эндообъект
122 0 объект
>
эндообъект
124 0 объект
[155 0 R 156 0 R 157 0 R 158 0 R 159 0 R 159 0 R 412 0 R 413 0 R 414 0 R 161 0 R 162 0 R]
эндообъект
125 0 объект
> эндообъект
126 0 объект
[163 0 Ч 164 0 Ч 165 0 Ч 166 0 Ч 167 0 Ч]
эндообъект
127 0 объект
[168 0 Р 1690 Р 170 0 Р 171 0 Р]
эндообъект
128 0 объект
[172 0 Р 173 0 Р 174 0 Р 175 0 Р 176 0 Р 177 0 Р]
эндообъект
129 0 объект
[178 0 Р 179 0 Р 181 0 Р 182 0 Р 180 0 Р]
эндообъект
130 0 объект
[183 0 Р 184 0 Р 185 0 Р 186 0 Р 187 0 Р]
эндообъект
131 0 объект
[188 0 Р 189 0 Р 190 0 Р]
эндообъект
132 0 объект
[191 0 Р 193 0 Р 192 0 Р]
эндообъект
133 0 объект
[194 0 Ч 195 0 Ч 196 0 Ч 197 0 Ч]
эндообъект
134 0 объект
[198 0 R 416 0 R 417 0 R 418 0 R 419 0 R 420 0 R 421 0 R 422 0 R 423 0 R 424 0 R 425 0 R 426 0 R 427 0 R 428 0 R 429 0 R 430 0 R 431 0 Р 432 0 Р 433 0 Р 434 0 Р 435 0 Р 436 0 Р 437 0 Р 438 0 Р 4390 R 440 0 R 441 0 R 442 0 R 443 0 R 444 0 R 445 0 R 446 0 R 447 0 R 448 0 R 449 0 R 450 0 R 451 0 R 452 0 R 453 0 R 454 0 R 455 0 R 456 0 Р 457 0 Р 458 0 Р 459 0 Р 460 0 Р 461 0 Р 462 0 Р 463 0 Р 464 0 Р 465 0 Р 466 0 Р 467 0 Р 468 0 Р 469 0 Р 470 0 Р 471 0 Р 472 0 Р 473 0 Р 474 0 Р 475 0 Р 476 0 Р 477 0 Р 478 0 Р 479 0 Р 480 0 Р 481 0 Р 482 0 Р 483 0 Р 484 0 Р 485 0 Р 486 0 Р 487 0 Р 488 0 Р 489 0 R 490 0 R 491 0 R 492 0 R 493 0 R 494 0 R 495 0 R 496 0 R 497 0 R 498 0 R 499 0 R 500 0 R 501 0 R 502 0 R 503 0 R 504 0 R 505 0 R 506 0 Р 507 0 Р 508 0 Р 5090 R 510 0 R 511 0 R 512 0 R 513 0 R 514 0 R 515 0 R 516 0 R 517 0 R 518 0 R 519 0 R 520 0 R 521 0 R 522 0 R 523 0 R 524 0 R 525 0 R 526 0 Р 527 0 Р 528 0 Р 529 0 Р 530 0 Р 531 0 Р 532 0 Р 533 0 Р 534 0 Р 535 0 Р 536 0 Р 537 0 Р 538 0 Р 539 0 Р 540 0 Р 541 0 Р 542 0 Р 543 0 Р 544 0 Р 545 0 Р 546 0 Р 547 0 Р 548 0 Р 549 0 Р 550 0 Р 551 0 Р 552 0 Р 553 0 Р 554 0 Р 555 0 Р 556 0 Р 557 0 Р 558 0 Р 559 0 R 560 0 R 561 0 R 562 0 R 563 0 R 564 0 R 565 0 R 566 0 R 567 0 R 568 0 R 569 0 R 570 0 R 571 0 R 572 0 R 573 0 R 574 0 R 575 0 R 576 0 R 577 0 R 200 0 R 201 0 R 202 0 R 203 0 R 204 0 R 205 0 R]
эндообъект
135 0 объект
[206 0 Р 208 0 Р 207 0 Р]
эндообъект
136 0 объект
[2090 R 210 0 R 211 0 R 212 0 R 214 0 R 215 0 R 216 0 R 213 0 R]
эндообъект
137 0 объект
[217 0 Р 218 0 Р 219 0 Р]
эндообъект
138 0 объект
[220 0 R 221 0 R 222 0 R 223 0 R 224 0 R 225 0 R 226 0 R]
эндообъект
139 0 объект
[227 0 Р 228 0 Р 229 0 Р 230 0 Р 231 0 Р]
эндообъект
140 0 объект
[232 0 Р 233 0 Р 234 0 Р 235 0 Р 236 0 Р 237 0 Р]
эндообъект
141 0 объект
[238 0 Ч 239 0 Ч 240 0 Ч 241 0 Ч 242 0 Ч]
эндообъект
142 0 объект
[243 0 Р 244 0 Р 245 0 Р 246 0 Р 247 0 Р 248 0 Р]
эндообъект
143 0 объект
[249 0 Р 250 0 Р 251 0 Р 252 0 Р 253 0 Р 254 0 Р]
эндообъект
144 0 объект
[255 0 Р 256 0 Р 257 0 Р 258 0 Р 2590 Р 260 0 Р 261 0 Р]
эндообъект
145 0 объект
[262 0 Р 263 0 Р 264 0 Р 265 0 Р 266 0 Р 267 0 Р 268 0 Р]
эндообъект
146 0 объект
[269 0 Р 270 0 Р 271 0 Р 272 0 Р 273 0 Р]
эндообъект
147 0 объект
[274 0 R 275 0 R 276 0 R 277 0 R 278 0 R 279 0 R 280 0 R 281 0 R 282 0 R 283 0 R 284 0 R 285 0 R 286 0 R 287 0 R 288 0 R 289 0 R 290 0 Р 291 0 Р 292 0 Р]
эндообъект
148 0 объект
[293 0 Р 294 0 Р 295 0 Р 296 0 Р 297 0 Р 298 0 Р 299 0 Р 300 0 Р 301 0 Р 302 0 Р 303 0 Р 304 0 Р 305 0 Р 306 0 Р 307 0 Р 308 0 Р 309 0 Р 310 0 Р 311 0 Р 312 0 Р 313 0 Р 314 0 Р 315 0 Р 316 0 Р 317 0 Р 318 0 Р 3190 Р 320 0 Р]
эндообъект
149 0 объект
[321 0 R 322 0 R 323 0 R 324 0 R 325 0 R 326 0 R 327 0 R 328 0 R 329 0 R 330 0 R 331 0 R 332 0 R 333 0 R 334 0 R 335 0 R 336 0 R 337 0 R 338 0 R 339 0 R 340 0 R 341 0 R 342 0 R 343 0 R 344 0 R 345 0 R]
эндообъект
150 0 объект
[346 0 Ч 347 0 Ч 348 0 Ч 350 0 Ч 351 0 Ч 349 0 Ч]
эндообъект
151 0 объект
[352 0 Ч 354 0 Ч 355 0 Ч 353 0 Ч]
эндообъект
152 0 объект
[356 0 R 358 0 R 359 0 R 360 0 R 361 0 R 362 0 R 363 0 R 364 0 R 365 0 R 366 0 R 367 0 R 368 0 R 369 0 R 370 0 R 371 0 R 357 0 R]
эндообъект
153 0 объект
[372 0 Р 373 0 Р 374 0 Р 375 0 Р 376 0 Р 377 0 Р 378 0 Р 3790 R 380 0 R 381 0 R 382 0 R 383 0 R 384 0 R 385 0 R 386 0 R 387 0 R 388 0 R 389 0 R 390 0 R 391 0 R 392 0 R 393 0 R 394 0 R]
эндообъект
154 0 объект
[395 0 Р 396 0 Р 397 0 Р 398 0 Р 399 0 Р 400 0 Р 401 0 Р 402 0 Р 403 0 Р 404 0 Р 405 0 Р 406 0 Р 407 0 Р 408 0 Р 409 0 Р 410 0 Р 411 0 Р]
эндообъект
395 0 объект
> эндообъект
396 0 объект
>
эндообъект
397 0 объект
>
эндообъект
398 0 объект
>
эндообъект
399 0 объект
>
эндообъект
400 0 объект
>
эндообъект
401 0 объект
>
эндообъект
402 0 объект
>
эндообъект
403 0 объект
>
эндообъект
404 0 объект
>
эндообъект
405 0 объект
>
эндообъект
406 0 объект
>
эндообъект
407 0 объект
>
эндообъект
408 0 объект
>
эндообъект
409mۑOc1p\x»R&»vl>spokenKy#;2h{_GѯcVfɗy&N2f .
 v9’PZrNy4*V4’4
BO#@zGg-f`Z9zئ:]:7Rqw3dk|WnsD?
gQHWU$z 6,#}ɕed ӶU#t*j`spoken 99ڴ2#K6
v9’PZrNy4*V4’4
BO#@zGg-f`Z9zئ:]:7Rqw3dk|WnsD?
gQHWU$z 6,#}ɕed ӶU#t*j`spoken 99ڴ2#K6FON4 предотвращает появление многоцветковых колосков у риса
- Список журналов
- Wiley-Blackwell Online Open
- PMC6524161
Журнал биотехнологии растений
Plant Biotechnol J. июнь 2019 г.; 17(6): 1007–1009.
Опубликовано в Интернете 6 февраля 2019 г. doi: 10.1111/pbi.13083
, 1 , † , 1 , † , 1 , † , 1 , 1 , 1 , 1 и 1 , 2Информация об авторе Примечания к статье Информация об авторских правах и лицензиях Отказ от ответственности
Колосок является особой единицей соцветия травы и дает различное количество цветков (1–40) в зависимости от того, демонстрирует ли вид детерминантный или индетерминантный рост.

В предыдущих исследованиях мутант fon4 продуцировал увеличенные внутренние цветочные органы и удлиненные стерильные цветковые цветковые чешуи с неопределенной идентичностью (Chu et al ., 2006), тогда как мы подтвердили, что 9Мутант 0059 fon4-7 показал два цветка и индуцировал реверсию стерильных цветковых чешуй в цветковые чешуи (боковые цветки). Мы обсуждаем две возможности формирования многоцветкового колоска у риса, которые могут способствовать выведению сортов риса с повышенным числом зерен на рисовой метелке.
У риса строго образуется пара стерильных цветковых чешуй и один цветочек на каждом колоске, каждый из которых содержит одну цветковую чешую и одну цветковую чешую, две лодыжки, примыкающие к цветовой чешуе, шесть тычинок и пестик (рис. A1–A5).
Открыть в отдельном окне
A, исследования фенотипов колосков дикого типа и fon4-7 мутанта. А1 и А2, колосок дикого типа. А3 — эпидермальная поверхность палео у дикого типа. А4 и А5, гистологические наблюдения колоска дикого типа. А6 и А7, фон4-7 колосок с дополнительным чешуевидным органом, двумя редуцированными палеями и четырьмя лодикулами. A8, эпидермальная поверхность редуцированной палео в A7. A9 и A10, гистологический анализ fon4-7 колосок с дополнительным чешуевидным органом, двумя вырожденными палеями и четырьмя лодикулами. А11 и А12, фон4-7 колосок с дополнительным чешуевидным органом и увеличенной палеей. A13, эпидермальная поверхность экстралеммоподобного органа в A12. А14 и А15, гистологический анализ колоска fon4-7 с дополнительным леммоподобным органом и увеличенной палеей. А16 и А17, фон4-7 колосок с удлиненной стерильной цветочной чешуей и увеличенной палеей. А18 — эпидермальная поверхность удлиненной стерильной чешуи А17. А19А20 — гистологический анализ колоска fon4-7 с удлиненной стерильной цветочной чешуей и увеличенной палео. B, экспрессия гена DL в диком типе и мутанте fon4-7 .
А4 и А5, гистологические наблюдения колоска дикого типа. А6 и А7, фон4-7 колосок с дополнительным чешуевидным органом, двумя редуцированными палеями и четырьмя лодикулами. A8, эпидермальная поверхность редуцированной палео в A7. A9 и A10, гистологический анализ fon4-7 колосок с дополнительным чешуевидным органом, двумя вырожденными палеями и четырьмя лодикулами. А11 и А12, фон4-7 колосок с дополнительным чешуевидным органом и увеличенной палеей. A13, эпидермальная поверхность экстралеммоподобного органа в A12. А14 и А15, гистологический анализ колоска fon4-7 с дополнительным леммоподобным органом и увеличенной палеей. А16 и А17, фон4-7 колосок с удлиненной стерильной цветочной чешуей и увеличенной палеей. А18 — эпидермальная поверхность удлиненной стерильной чешуи А17. А19А20 — гистологический анализ колоска fon4-7 с удлиненной стерильной цветочной чешуей и увеличенной палео. B, экспрессия гена DL в диком типе и мутанте fon4-7 . B1-B4, колосок дикого типа. В1, Сп4; B2, Sp5-6; В3, Сп7, В4, Сп8. B5-B8, fon4-7 колосок с дополнительным чешуевидным органом и двумя редуцированными палеями. В5, Сп4; B6, Sp5-6; В7, Сп7, В8, Сп8. B9-B12, fon4-7 колосок с дополнительным чешуевидным органом и увеличенной палеей. В9, Сп4; B10, Sp5-6; В11, Сп7, В12, Сп8. B13-B16, fon4-7
B1-B4, колосок дикого типа. В1, Сп4; B2, Sp5-6; В3, Сп7, В4, Сп8. B5-B8, fon4-7 колосок с дополнительным чешуевидным органом и двумя редуцированными палеями. В5, Сп4; B6, Sp5-6; В7, Сп7, В8, Сп8. B9-B12, fon4-7 колосок с дополнительным чешуевидным органом и увеличенной палеей. В9, Сп4; B10, Sp5-6; В11, Сп7, В12, Сп8. B13-B16, fon4-7
 D, модель гипотетического разведения молекулярного дизайна и предполагаемой функции FON4 . В модели LF1 индуцирует боковые цветки в одиночном колоске риса с нормальной стерильной чешуей; ФОН4 , ТОБ1 , М34 , СНБ , Г1 , МФС1 , EG1/DF1 подавляют образование чешуевидных стерильных цветковых чешуй или дополнительных цветковых чешуй или двухцветковых колосков в одиночном колоске риса с нормальной стерильной чешуей или без стерильной чешуи; M22 индуцирует двухцветковый колосок в одиночном колоске риса с нормальной стерильной чешуей. esl – удлиненная стерильная цветковая чешуя; ell — экстралеммоподобный орган; ле, лемма; вот, лодикула; ст, тычинка; пи, пестик; па, палеа; pal, палеоподобный орган; rg — рудиментарная чешуя; sl — стерильная цветковая чешуя; ep, увеличенная палео-окраска; mrp — краевые области палео; боп, тело палео; о, внешне крючковидная структура; ih — крючковидное строение внутрь, fm — меристема цветка; W-p, метелка дикого типа; M-p, метелка с многоцветковым колоском; Т-р, метелка с двуцветковым колоском; W-s, колосок дикого типа; Mfs, многоцветковый колосок; Tfs-I, двухцветковый колосок без стерильной цветковой чешуи; Tfs-II, двухцветковый колосок со стерильной чешуей; М22 , ОсМАДС22 ; М34 , ОсМАДС34 .
D, модель гипотетического разведения молекулярного дизайна и предполагаемой функции FON4 . В модели LF1 индуцирует боковые цветки в одиночном колоске риса с нормальной стерильной чешуей; ФОН4 , ТОБ1 , М34 , СНБ , Г1 , МФС1 , EG1/DF1 подавляют образование чешуевидных стерильных цветковых чешуй или дополнительных цветковых чешуй или двухцветковых колосков в одиночном колоске риса с нормальной стерильной чешуей или без стерильной чешуи; M22 индуцирует двухцветковый колосок в одиночном колоске риса с нормальной стерильной чешуей. esl – удлиненная стерильная цветковая чешуя; ell — экстралеммоподобный орган; ле, лемма; вот, лодикула; ст, тычинка; пи, пестик; па, палеа; pal, палеоподобный орган; rg — рудиментарная чешуя; sl — стерильная цветковая чешуя; ep, увеличенная палео-окраска; mrp — краевые области палео; боп, тело палео; о, внешне крючковидная структура; ih — крючковидное строение внутрь, fm — меристема цветка; W-p, метелка дикого типа; M-p, метелка с многоцветковым колоском; Т-р, метелка с двуцветковым колоском; W-s, колосок дикого типа; Mfs, многоцветковый колосок; Tfs-I, двухцветковый колосок без стерильной цветковой чешуи; Tfs-II, двухцветковый колосок со стерильной чешуей; М22 , ОсМАДС22 ; М34 , ОсМАДС34 . Черные стрелки обозначают mrp в A14 и A19. Желтые стрелки и красные звезды обозначают сосудистые пучки в A. N указывает номер цветка в метелке в D. «X» в D указывает на скрещивание в разведении. Стержни = 1000 мм в форматах A1, A2, A6, A7, A11, A12, A16 и A17; 100 мкм в A3–A5, A8–A10, A13–A15 и A18–A20; и 50 мкм в B и C.
Черные стрелки обозначают mrp в A14 и A19. Желтые стрелки и красные звезды обозначают сосудистые пучки в A. N указывает номер цветка в метелке в D. «X» в D указывает на скрещивание в разведении. Стержни = 1000 мм в форматах A1, A2, A6, A7, A11, A12, A16 и A17; 100 мкм в A3–A5, A8–A10, A13–A15 и A18–A20; и 50 мкм в B и C. Колоски fon4-7 группы I развили дополнительные леммоподобные органы и два палеоподобных органа (рис. A5–A10). В этих колосках наблюдались четыре лодикулы, по две с каждой стороны каждой цветковой чешуи (Рис. A7–A9).). Этот фенотип указывал на то, что эти колоски образовали два цветка. Колоски группы II продуцировали вторичный цветочек, содержащий похожий на чешуйку орган (Рисунок A11–A15), тогда как колоски группы III образовали конечный цветочек с чешуевидной стерильной чешуей (боковые цветочки) (Рисунок A16–A20). Наши подробные фенотипические наблюдения показали, что все чешуйкоподобные органы, чешуйкоподобные стерильные чешуи и увеличенное тело палеаи имели такие же клеточные структуры и количество сосудистых пучков, что и чешуи дикого типа (рис. A4, A14, A19).). OsMADS1 , OsMADS14 и OsMADS15 экспрессировались в типичных чешуйках, дополнительных чешуевидных органах, удлиненных стерильных чешуях и бледноподобных органах мутанта fon4-7 . Экспрессия DL также была обнаружена в вышеуказанных органах, за исключением palea-подобных органов у мутанта fon4-7 . OsMADS6 экспрессировался в fon4-7 Palae и палеоподобных органах. Эти изменения в экспрессии показали, что дополнительные чешуйкоподобные органы и удлиненные стерильные чешуи fon4-7 колоски имели идентичность цветковых чешуй, а палеоподобный орган образовался из деградировавших палео. Повышенная экспрессия OsMADS2 и OsMADS6 у мутанта fon4-7 может быть связана с относительным обилием соответствующих органов у этого мутанта.
A4, A14, A19).). OsMADS1 , OsMADS14 и OsMADS15 экспрессировались в типичных чешуйках, дополнительных чешуевидных органах, удлиненных стерильных чешуях и бледноподобных органах мутанта fon4-7 . Экспрессия DL также была обнаружена в вышеуказанных органах, за исключением palea-подобных органов у мутанта fon4-7 . OsMADS6 экспрессировался в fon4-7 Palae и палеоподобных органах. Эти изменения в экспрессии показали, что дополнительные чешуйкоподобные органы и удлиненные стерильные чешуи fon4-7 колоски имели идентичность цветковых чешуй, а палеоподобный орган образовался из деградировавших палео. Повышенная экспрессия OsMADS2 и OsMADS6 у мутанта fon4-7 может быть связана с относительным обилием соответствующих органов у этого мутанта.
Мы наблюдали за развитием ранних колосков. На 4-й стадии колоска (Sp4) некоторые из колосков fon4-7 продуцировали дополнительные чешуевидные зачатки и широкие или узкие зачатки палео. На этапах Sp5 и Sp6 fon4-7 колоски образовывали большее количество тычинок и поддерживали рост их дефектной палеи. На Sp7 и Sp8 в колоске fon4-7 явно наблюдались дополнительные чешуевидные, палеоподобные органы и удлинённые стерильные чешуи, а лепестки были шире и имели более одной бугорчатой вершины. Явных различий стерильных цветковых чешуй между диким типом и мутантом fon4-7 на стадиях Sp4–Sp7 не обнаружено. На Sp8 стерильные чешуи fon4‐7 мутант резко дифференцировался и был сопоставим по размеру с нижними цветковыми чешуями дикого типа.
На этапах Sp5 и Sp6 fon4-7 колоски образовывали большее количество тычинок и поддерживали рост их дефектной палеи. На Sp7 и Sp8 в колоске fon4-7 явно наблюдались дополнительные чешуевидные, палеоподобные органы и удлинённые стерильные чешуи, а лепестки были шире и имели более одной бугорчатой вершины. Явных различий стерильных цветковых чешуй между диким типом и мутантом fon4-7 на стадиях Sp4–Sp7 не обнаружено. На Sp8 стерильные чешуи fon4‐7 мутант резко дифференцировался и был сопоставим по размеру с нижними цветковыми чешуями дикого типа.
Затем мы исследовали паттерны экспрессии генов идентичности органов цветка с использованием гибридизации in situ . У дикого типа экспрессия DL была обнаружена в цветковой чешуе на стадиях от Sp4 до Sp7 и в средней жилке цветковой чешуи на стадии Sp8 (рис. B1–B4). У мутанта fon4-7 экспрессия DL была обнаружена в нормальных цветковых чешуях, дополнительных леммоподобных органах и удлиненных стерильных цветковых чешуях (рис. B5–B16). Эти результаты подтвердили, что мутантные колоски действительно образовывали дополнительную чешуйку или подвергались гомологичному превращению стерильной чешуи в чешуйку. На этапах от Sp4 до Sp7 Экспрессия OsMADS6 у мутанта fon4-7 напоминала экспрессию дикого типа, при этом сигналы транскрипции наблюдались в меристеме цветков, краевых областях палео (mrp), лодикулах и пестиках (рис. C1–C3, C5–C7, С9–С11, С11–С15). В Sp8 fon4-7 экспрессировали этот ген в четырех mrps двух палеоподобных органов (C4, C8, C12, C16). Эти находки также показали, что две палеоподобные структуры в одном колоске fon4-7 произошли от нормальных палеоподобных клеток. 9Сигналы 0059 OsMADS2 были обнаружены в тычинках и лодикулах, прилегающих к цветковой чешуе у дикого типа на стадиях Sp4–Sp8; однако в группе I fon4-7 колосков транскрипты этого гена были обнаружены в типичных лодикулах и экстралодикулах на стадиях от Sp4 до Sp7. На Sp8 сигналы транскрипции OsMADS2 были ярко выражены в двух лодикулах с каждой стороны цветковой чешуи в индивидуальном колоске fon4-7 , что подтверждает, что четыре лодикулы были произведены в одиночном фон4‐7 колосок, содержащий две цветковые чешуи.
B5–B16). Эти результаты подтвердили, что мутантные колоски действительно образовывали дополнительную чешуйку или подвергались гомологичному превращению стерильной чешуи в чешуйку. На этапах от Sp4 до Sp7 Экспрессия OsMADS6 у мутанта fon4-7 напоминала экспрессию дикого типа, при этом сигналы транскрипции наблюдались в меристеме цветков, краевых областях палео (mrp), лодикулах и пестиках (рис. C1–C3, C5–C7, С9–С11, С11–С15). В Sp8 fon4-7 экспрессировали этот ген в четырех mrps двух палеоподобных органов (C4, C8, C12, C16). Эти находки также показали, что две палеоподобные структуры в одном колоске fon4-7 произошли от нормальных палеоподобных клеток. 9Сигналы 0059 OsMADS2 были обнаружены в тычинках и лодикулах, прилегающих к цветковой чешуе у дикого типа на стадиях Sp4–Sp8; однако в группе I fon4-7 колосков транскрипты этого гена были обнаружены в типичных лодикулах и экстралодикулах на стадиях от Sp4 до Sp7. На Sp8 сигналы транскрипции OsMADS2 были ярко выражены в двух лодикулах с каждой стороны цветковой чешуи в индивидуальном колоске fon4-7 , что подтверждает, что четыре лодикулы были произведены в одиночном фон4‐7 колосок, содержащий две цветковые чешуи. Эти результаты подтверждают вывод о том, что в одном колоске fon4-7 образовались два независимых цветка.
Эти результаты подтверждают вывод о том, что в одном колоске fon4-7 образовались два независимых цветка.
Локус FON4 был сужен между маркерами М7 и М13. Секвенирование ДНК привело к идентификации аминокислотной мутации в мутанте fon4-7 внутри Os11g38270 . Анализ комплементации показал, что фенотипов fon4-7 были полностью восстановлены. CRISPR/Cas9выбитые мутанты fon4-8 и fon4-9 демонстрировали фенотипы, которые напоминали мутант fon4-7 . Эти результаты дополнительно подтвердили, что FON4 представляет собой ген Os11g38270 . Дальнейшие исследования показали, что fon4-8 и fon4-9 индуцируют вариабельное количество многоцветковых колосков, предполагая, что различные сайты вариации или генетический фон в гене FON4 привели к большему и меньшему количеству многоцветковых колосков.
Около 41% из fon4-7 колосков образовали типичную цветковую чешую и дополнительную цветковую чешую, два палеоподобных органа, увеличенные тычинки и пестики и четыре лодыжки, в которых по две с каждой стороны каждой цветковой чешуи, предполагая, что два цветка формируются. Около 16% из fon4-7 колосков несли нормальные цветки с дополнительными цветковыми чешуями, что означало, что эти колоски состояли из верхушечного цветка и вторичного цветка, содержащего только чешуйку. Эти данные позволяют предположить, что детерминированность меристемы колоска была утрачена в 9-м веке.0059 fon4-7 мутант. Этот фенотип с двумя цветками или дополнительным леммоподобным органом (вторичный цветочек) также наблюдался ранее у мутантов tob1 , mfs1 , snb и eg1 / df1 , а также у СНБ + osids1 двойной мутант и OsMADS22 -сверхэкспрессирующие растения (Lee et al ., 2007; Li et al ., 2009; Ren et al ., 2013, 2018; Сентоку и др. ., 2005; Танака и др. ., 2012). Недавнее исследование показало, что мутация LF1 приводит к образованию одного или двух фертильных боковых цветков (Zhang et al ., 2017). Эти исследования позволяют предположить, что FON4 , LF1 , TOB1 , MFS1 , SNB и OsMADS22 регулируют детерминированность меристемы колоска или их мутации.
Около 16% из fon4-7 колосков несли нормальные цветки с дополнительными цветковыми чешуями, что означало, что эти колоски состояли из верхушечного цветка и вторичного цветка, содержащего только чешуйку. Эти данные позволяют предположить, что детерминированность меристемы колоска была утрачена в 9-м веке.0059 fon4-7 мутант. Этот фенотип с двумя цветками или дополнительным леммоподобным органом (вторичный цветочек) также наблюдался ранее у мутантов tob1 , mfs1 , snb и eg1 / df1 , а также у СНБ + osids1 двойной мутант и OsMADS22 -сверхэкспрессирующие растения (Lee et al ., 2007; Li et al ., 2009; Ren et al ., 2013, 2018; Сентоку и др. ., 2005; Танака и др. ., 2012). Недавнее исследование показало, что мутация LF1 приводит к образованию одного или двух фертильных боковых цветков (Zhang et al ., 2017). Эти исследования позволяют предположить, что FON4 , LF1 , TOB1 , MFS1 , SNB и OsMADS22 регулируют детерминированность меристемы колоска или их мутации.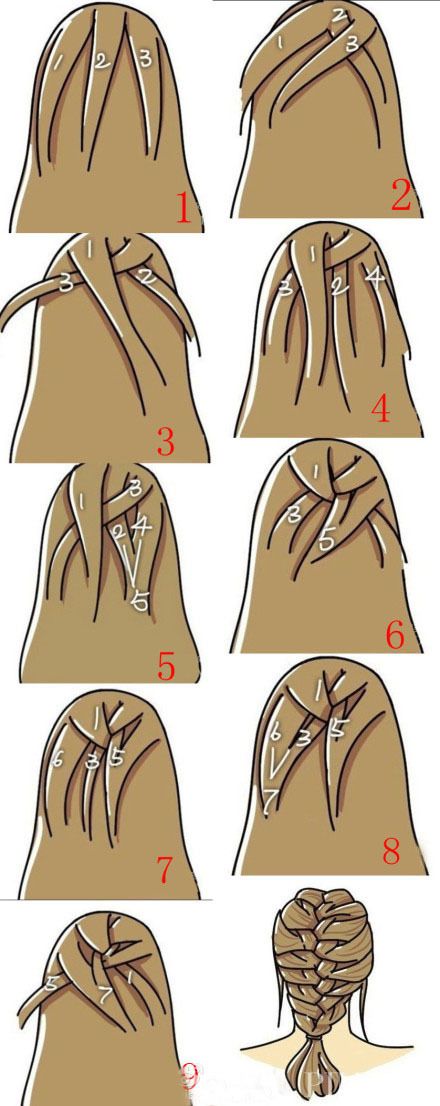 или эктопические экспрессии индуцируют неопределенность меристемы колоска. С пшеницей и овсом ( Avena sativa ), меристема колоска проявляет индетерминантность и формирует множественные цветки, образуя более одного семени в колоске. Эти результаты показывают, что можно получить два или более соцветия в каждом отдельном колоске риса, что повышает вероятность увеличения количества семян, производимых на одно растение. Кроме того, мы обнаружили, что мутанты fon4 продуцируют различное количество многоцветковых колосков, что указывает на то, что мутация различных нуклеотидов в FON4 приводит к различному влиянию на его способность контролировать детерминированность колосков, тем самым производя больше и меньше мультицветков. -цветковый колосок.
или эктопические экспрессии индуцируют неопределенность меристемы колоска. С пшеницей и овсом ( Avena sativa ), меристема колоска проявляет индетерминантность и формирует множественные цветки, образуя более одного семени в колоске. Эти результаты показывают, что можно получить два или более соцветия в каждом отдельном колоске риса, что повышает вероятность увеличения количества семян, производимых на одно растение. Кроме того, мы обнаружили, что мутанты fon4 продуцируют различное количество многоцветковых колосков, что указывает на то, что мутация различных нуклеотидов в FON4 приводит к различному влиянию на его способность контролировать детерминированность колосков, тем самым производя больше и меньше мультицветков. -цветковый колосок.
Предыдущие исследования показали, что мутация G1 , DF1 / EG1 или OsMADS34 вызывает образование чешуевидных стерильных чешуй, тогда как мутация LF1 индуцирует образование боковых цветков без цветковых чешуй (Li et al . , 2009; Lin et al ., 2014; Yoshida et al ., 2009; Zhang et al ., 2017). Эти морфологические находки убедительно подтвердили гипотезу трехцветкового колоска (Zhang и др. ., 2017). Утрата функций FON4 , DF1 / EG1 , TOB1 , MFS1 и SNB вызывает развитие экстрачешуевидного органа или образование двух цветков. ets внутри одного колоска, что потенциально позволяет развитие остаточных меристем колоска или превращение детерминированности в индетерминантность. Эти данные дают две возможности для селекции сортов риса с многоцветковым колоском. Один из вариантов — пересечь lf1 мутант с нашим мутантом fon4 или другими мутантами ( m34 , g1 , mfs1 и eg1 ), которые производят леммоподобные стерильные цветковая чешуя или дополнительный чешуевидный орган в пределах одного колоска, с целью в конечном итоге создать сорта с двух- или трехцветковым колоском (рис.
, 2009; Lin et al ., 2014; Yoshida et al ., 2009; Zhang et al ., 2017). Эти морфологические находки убедительно подтвердили гипотезу трехцветкового колоска (Zhang и др. ., 2017). Утрата функций FON4 , DF1 / EG1 , TOB1 , MFS1 и SNB вызывает развитие экстрачешуевидного органа или образование двух цветков. ets внутри одного колоска, что потенциально позволяет развитие остаточных меристем колоска или превращение детерминированности в индетерминантность. Эти данные дают две возможности для селекции сортов риса с многоцветковым колоском. Один из вариантов — пересечь lf1 мутант с нашим мутантом fon4 или другими мутантами ( m34 , g1 , mfs1 и eg1 ), которые производят леммоподобные стерильные цветковая чешуя или дополнительный чешуевидный орган в пределах одного колоска, с целью в конечном итоге создать сорта с двух- или трехцветковым колоском (рис. D). Другая возможность состоит в том, чтобы идентифицировать различные сайты-мишени для редактирования генов в генах ( FON4 , DF1 / EG1 , TOB1 9).0060 , MFS1 , SNB и M22 ), участвующие в регуляции детерминированности и индетерминантности меристемы колосков (рис. D). Колосок с несколькими цветками может увеличить количество зерен на метелке, тем самым увеличивая урожайность риса.
D). Другая возможность состоит в том, чтобы идентифицировать различные сайты-мишени для редактирования генов в генах ( FON4 , DF1 / EG1 , TOB1 9).0060 , MFS1 , SNB и M22 ), участвующие в регуляции детерминированности и индетерминантности меристемы колосков (рис. D). Колосок с несколькими цветками может увеличить количество зерен на метелке, тем самым увеличивая урожайность риса.
Эта работа была поддержана Фондом естественных наук Чжэцзяна (LY18C130007), Центральным общественным научным учреждением, Фондом фундаментальных исследований Китайского национального исследовательского института риса (2017RG001-4), Национальным фондом естественных наук Китая (31521064 и 9).1735304), Национальный крупный проект по науке и технологиям (2016ZX08009003-003-008) и Национальная программа селекции новых сортов ГМО КНР (2016ZX08011-001).
Авторы заявляют, что у них нет конкурирующих интересов.
Дейонг Рен, электронная почта: moc. 361@616gnoyedner.
361@616gnoyedner.
Цянь Цянь, электронная почта: moc.liamtoh@881naiqnaiq.
- Чу, Х.В. , Цянь, К. , Лян, В.К. , Инь, К.С. , Тан, Х.Х. , Яо, X. , Юань, З. и др. (2006) НОМЕР ЦВЕТОЧНОГО ОРГАНА4 9Ген 0060, кодирующий предполагаемый ортолог Arabidopsis CLAVATA3, регулирует размер апикальной меристемы риса. Завод Физиол. 142, 1039–1052. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Ли, Д.Ю. , Ли, Дж. , Мун, С. , Парк, С.Ю. и Ан, Г. (2007) Ген гетерохронности риса SUPERNUMERARY BRACT регулирует переход от меристемы колоска к меристеме цветка. Плант Дж. 49, 64–78. [PubMed] [Академия Google]
- Ли, Х.Г. , Сюэ, Д.В. , Гао, З.Ю. , Ян, М.Х. , Сюй, В.Ю. , Син, З. , Хуанг, Д.Н. et al (2009) Предполагаемый ген липазы EXTRA GLUME1 регулирует как судьбу пустых чешуй, так и развитие колосков у риса. Плант Дж. 57, 593–605. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Лин, С.Л.
,
Ву, Ф.
 ,
Ду, X.Q.
,
Ши, X.W.
,
Лю, Ю.
,
Лю, С.Дж.
,
Ху, Ю.С. и др. (2014). Плейотропный SEPALLATA-подобный ген OsMADS34 показывает, что «пустые чешуйки» риса ( Oryza sativa ) колосков на самом деле являются рудиментарными цветковыми чешуями. Новый Фитол.
202, 689–702. [PubMed] [Академия Google]
,
Ду, X.Q.
,
Ши, X.W.
,
Лю, Ю.
,
Лю, С.Дж.
,
Ху, Ю.С. и др. (2014). Плейотропный SEPALLATA-подобный ген OsMADS34 показывает, что «пустые чешуйки» риса ( Oryza sativa ) колосков на самом деле являются рудиментарными цветковыми чешуями. Новый Фитол.
202, 689–702. [PubMed] [Академия Google] - Рен, Д.Ю. , Ли, Ю.Ф. , Чжао, Ф.Н. , Санг, X.C. , Ши, Дж.К. , Ван, Н. , Го, С. и др. (2013 г.) MULTI-FLORET SPIKELET1 , который кодирует белок AP2/ERF, определяет судьбу меристемы колоска и идентичность стерильной цветочной чешуи у риса. Завод Физиол. 162, 872–884. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Рен, Д.Ю. , Ю, Х.П. , Рао, Ю.К. , Сюй, К.К. , Чжоу, Т.Т. , Ху, Дж. , Чжан, Ю. et al (2018) «Двухцветковый колосок» как новый ресурс может повысить урожайность риса. Биотехнология растений. Дж. 16, 351–353. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Сентоку, Н.
,
Хидэки, Като.
,
Китано, Х. и
Имай, Р.
(2005) OsMADS22 , STMADS11-подобный ген MADS-box риса, экспрессируется в невегетативных тканях, и его эктопическая экспрессия индуцирует неопределенность меристемы колосков. Мол. Жене. Геномика, 273, 1–9. [PubMed] [Академия Google]
- Танака, В. , Ториба, Т. , Омори, Ю. , Йошида, А. , Кавай, А. , Маяма-Цучида, Т. , Итикава, Х. et al (2012). Ген YABBY TONGARI-BOUSHI1 участвует в развитии латеральных органов и поддержании меристеморганизации в колоске риса. Растительная клетка, 24, 80–9.5. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Йошида, А. , Сузаки, Т. , Танака, В. и Хирано, Х. (2009) Гомеозисный ген long sterile lemma ( G1 ) определяет идентичность стерильной леммы в колоске риса. проц. Натл акад. науч. США, 106, 20103–20108 гг. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Чжан, Т.
,
Ли, Ю.Ф.
,
Ма, Л.
,
Санг, X.C.
,
Линг, Ю.Х.
,
Ван, Ю.Т.
,
Ага. и др. (2017 г.) БОКОВОЙ ЦВЕТОК 1 индуцировал образование трехцветкового колоска у риса.
 ,
Ду, X.Q.
,
Ши, X.W.
,
Лю, Ю.
,
Лю, С.Дж.
,
Ху, Ю.С. и др. (2014). Плейотропный SEPALLATA-подобный ген OsMADS34 показывает, что «пустые чешуйки» риса ( Oryza sativa ) колосков на самом деле являются рудиментарными цветковыми чешуями. Новый Фитол.
202, 689–702. [PubMed] [Академия Google]
,
Ду, X.Q.
,
Ши, X.W.
,
Лю, Ю.
,
Лю, С.Дж.
,
Ху, Ю.С. и др. (2014). Плейотропный SEPALLATA-подобный ген OsMADS34 показывает, что «пустые чешуйки» риса ( Oryza sativa ) колосков на самом деле являются рудиментарными цветковыми чешуями. Новый Фитол.
202, 689–702. [PubMed] [Академия Google]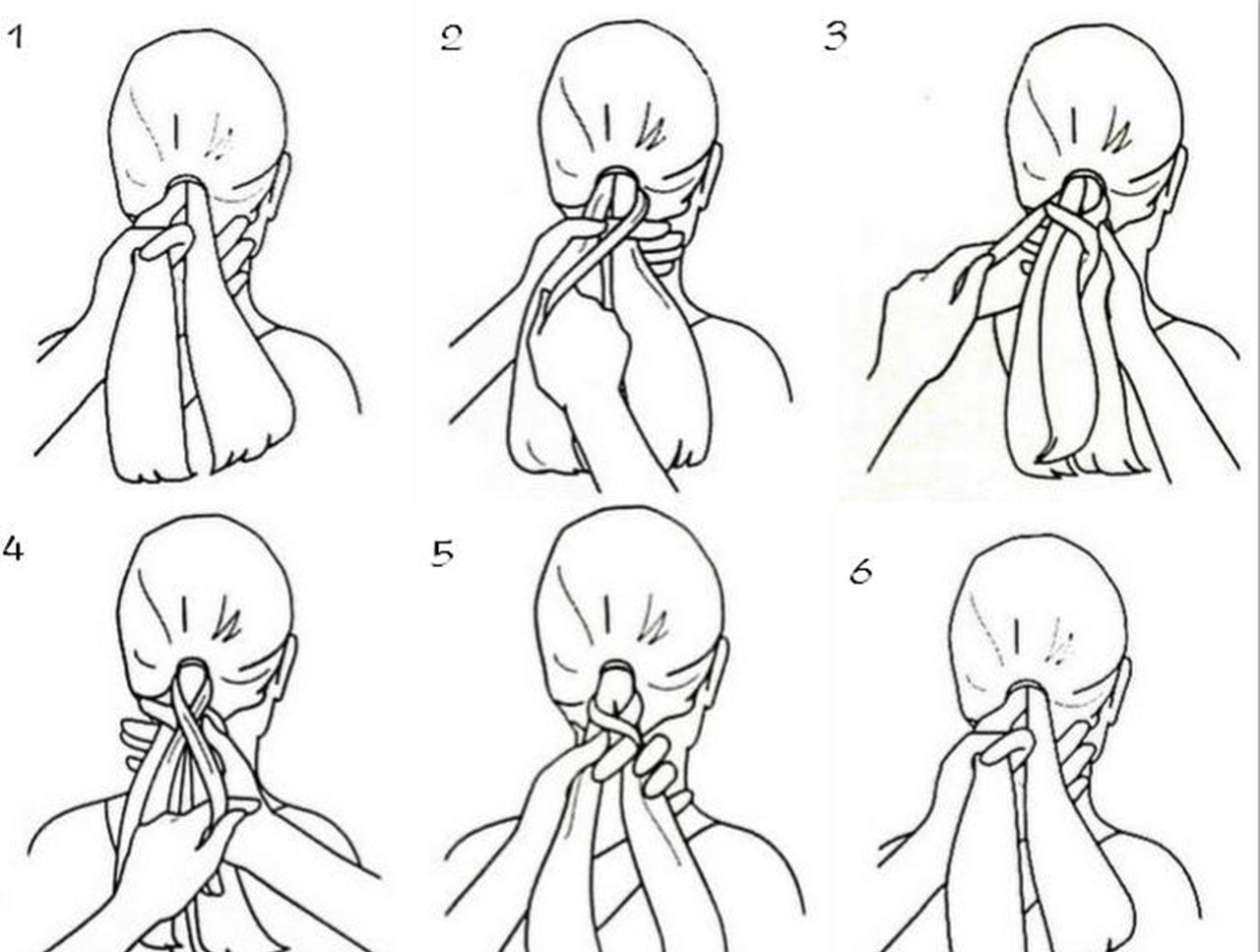 и
Имай, Р.
(2005) OsMADS22 , STMADS11-подобный ген MADS-box риса, экспрессируется в невегетативных тканях, и его эктопическая экспрессия индуцирует неопределенность меристемы колосков. Мол. Жене. Геномика, 273, 1–9. [PubMed] [Академия Google]
и
Имай, Р.
(2005) OsMADS22 , STMADS11-подобный ген MADS-box риса, экспрессируется в невегетативных тканях, и его эктопическая экспрессия индуцирует неопределенность меристемы колосков. Мол. Жене. Геномика, 273, 1–9. [PubMed] [Академия Google]