Как заплести косу? 5 видов плетения косичек
Косички – это универсальная прическа, которая подойдет как для строгой обстановки, так и для праздничного вечера. Сейчас причесок с косичками такое разнообразие, что их можно подобрать на любой вкус и волосы разной длины. Такая прическа подчеркнет силуэт, а волосы не будут мешаться при работе.
Обычная коса
Традиционная коса плетется в три пряди. Все волосы перекидываются назад, делятся на три одинаковые части и заплетаются в косу, соблюдая последовательность: вначале третью прядь перекидывают на первую и вторую прядь, затем первую на вторую и третью, и вторую прядь на третью и первую. Важно следить, чтобы волосы не выбивались, и стягивать их потуже, так заплетается коса и теперь ее можно закрепить красивой резинкой или лентой.
Колосок
Плетение колоска несколько более сложно, чем плетение косы, но техники у них очень схожи. Первый этап плетения колоска такой же, как у обычной косы – волосы зачесываются назад и, начиная от затылка следует выделить три небольшие пряди. Толщина прядей будет потом влиять на размер косы, но обычно классическую прядь выделяют примерно размером 2,5 сантиметров.
Толщина прядей будет потом влиять на размер косы, но обычно классическую прядь выделяют примерно размером 2,5 сантиметров.
Размер каждой пряди должен быть одинаковым, иначе колосок получится не ровным. Теперь эти пряди плетутся, как обычная коса. Следующий этап прически – это вплетение в косу новых прядей небольшого размера из боков. Это делается следующим образом – большим пальцем правой руки захватывай от низа прядь небольшого размера и вплетай в правую прядь основы, потом начинай опять заплетать косу, как обычно.
Далее также захвати левой рукой прядь волос из низа и вплетай к основной левой пряди, увеличенную в размере прядь также вплетай в косу. Таким образом, продолжай вплетать боковые пряди в основную косу, пока не закончится линия роста волос на голове. Если колосок получился недостаточно объемный, то можно придать ему густоту расческой с тонкими зубьями слегка распушив его. Ну и самый последний этап – это закрепить остаток волос, сделав из него хвост или доплести обычную косу до конца.
Секреты ровного колоска
Правильно заплетенный колосок никогда не подведет и не распадется на мелкие пряди в течение всего дня. Но часто новички в заплетании колоска сталкиваются с проблемой выбивания мелких прядей из косы. Чтобы избежать этого, следует выбирать пряди снизу строго по направлению плетения и как можно туже их натягивать. Прическа продержится очень долго в отличном состоянии, если колосок будет заплетен максимально туго.
Также можно применять для такой прически средства для укладки, чтобы волосы становились послушными – к примеру, некоторые неровные пряди можно смазать пенкой или закрепить заколками, так прическа будет смотреться более гладкой и ухоженной.
Есть еще и виды плетений, где для косы выделяются четыре или пять прядей, много вариаций французской косы, где плетение начинается не с затылка, а теменной области.
Французская коса наизнанку
Французская коса наоборот плетется точно также, как и обычное плетение косы, но при этом пряди вплетаются под центральную прядь, а не на нее. Давай разберем подробно, как это делается. Вначале собери все волосы с макушки, лба и боков и сделай из них пучок, а теперь раздели его на три ровных части. Начинай заплетать обычную косу, но пряди заплетай не поверх плетения, а под него. Должно получиться так, что заплетенные локоны будут обращены изнанкой наружу. При втором переплетении прядей, возьми из одной стороны другую часть волос и прибавь ее к общей части косички. Продолжай заплетать пряди до затылка, а теперь начинай заплетать обычную косу, которая будет обращена вершиной к низу. Особенность этой французской косы в том, что она более выпуклая и заметная, но это уже как говорится дело вкуса.
Давай разберем подробно, как это делается. Вначале собери все волосы с макушки, лба и боков и сделай из них пучок, а теперь раздели его на три ровных части. Начинай заплетать обычную косу, но пряди заплетай не поверх плетения, а под него. Должно получиться так, что заплетенные локоны будут обращены изнанкой наружу. При втором переплетении прядей, возьми из одной стороны другую часть волос и прибавь ее к общей части косички. Продолжай заплетать пряди до затылка, а теперь начинай заплетать обычную косу, которая будет обращена вершиной к низу. Особенность этой французской косы в том, что она более выпуклая и заметная, но это уже как говорится дело вкуса.
Плетение в виде «ободка»
Иногда хочется похвастаться своими длинными роскошными волосами, но при этом добавить прическе немного оригинальности. В таком случае подойдет небольшая косичка ленточкой или ободком. Обладательницы кучерявых и непослушных волос могут быть благодарны такой прическе – она позволит им забыть о без конца падающих и мешающихся прядях и при этом подчеркнет красоту и богатство их волос. Чтобы заплести такую косу делаем пробор посредине головы, проводя его от одного уха к другому, а ото лба надо сделать отступ примерно в 7 сантиметров.
Чтобы заплести такую косу делаем пробор посредине головы, проводя его от одного уха к другому, а ото лба надо сделать отступ примерно в 7 сантиметров.
Спереди волосы плетем французской или обычной косой, начиная плетение по лбу от уха к уху. Затем кончик косы прикрепляем шпилькой или заколкой за волосами, чтобы его не было видно, оставшуюся часть волос взбиваем руками, придаем объем с помощью начеса и заливаем волосы лаком.
Косичка из пяти прядей
Для этой косы тебе потребуются зажимы или помощница, чтобы придерживать пряди. Волосы слегка увлажни и зачеши их назад. Теперь раздели их на пять равных прядей и правую прядь скрести с ближайшей к ней прядью.
Прядь, которая самая центральная скрести с прядью, которая до этого была правая. Затем возьми вновь центральную прядь, и скрести с прядью, которая располагается левее, но не самая крайняя. Самую левую прядь теперь скрести с соседней прядью с правой стороны. Выполняй все эти действия, пока не заплетешь косу до конца.
Читайте также:
- Плетение кос на средние волосы
- Плетение косичек: схемы
- Прическа Французский водопад: схема плетения
- Как плести рыбий хвост?
- Как научиться красиво плести косы?
- Как заплести французкую косу?
- Из 5 прядей коса: схемы плетения
Radio Weave Cane Webbing шириной 18 дюймов, 5 нитей на дюйм
- Нажмите, чтобы увеличить
В 8 тележках
Цена: 279.00 турецких лир
Загрузка
Включены местные налоги (где применимо)
Принятые возвраты
Количество12345678910111213141516171819202122232425262728293031323334353637383940414243444546474849505152535455565758596 061626364656667686970717273747576777879808182838485868788899091929394959697989910010110210310410510610710810911011111211311 41151161171181191201211221231241251261271281291301311321333134135136137138139140141142143144414514614714814915015115215315415 5156157158159160161162163164165166167168169170171172173
Вы можете сделать предложение только при покупке одного товара
Внесен в список 7 июня 2023 г.
619 избранных
Сообщить об этом элементе в Etsy
Выберите причину… С моим заказом возникла проблемаОн использует мою интеллектуальную собственность без разрешенияЯ не думаю, что это соответствует политике EtsyВыберите причину…
Первое, что вы должны сделать, это связаться с продавцом напрямую.
Если вы уже это сделали, ваш товар не прибыл или не соответствует описанию, вы можете сообщить об этом Etsy, открыв кейс.
Сообщить о проблеме с заказом
Мы очень серьезно относимся к вопросам интеллектуальной собственности, но многие из этих проблем могут быть решены непосредственно заинтересованными сторонами. Мы рекомендуем связаться с продавцом напрямую, чтобы уважительно поделиться своими проблемами.
Если вы хотите подать заявление о нарушении прав, вам необходимо выполнить процедуру, описанную в нашей Политике в отношении авторских прав и интеллектуальной собственности.
Посмотрите, как мы определяем ручную работу, винтаж и расходные материалы
Посмотреть список запрещенных предметов и материалов
Ознакомьтесь с нашей политикой в отношении контента для взрослых
Товар на продажу…не ручной работы
не винтаж (20+ лет)
не ремесленные принадлежности
запрещены или используют запрещенные материалы
неправильно помечен как содержимое для взрослых
Пожалуйста, выберите причину
Расскажите нам больше о том, как этот элемент нарушает наши правила. Расскажите нам больше о том, как этот элемент нарушает наши правила. Все категории Товары для рукоделия и инструменты
Все категории Товары для рукоделия и инструментыДни плетения синтеза отстающих цепей ДНК. Личные воспоминания об открытии фрагментов Окадзаки и исследованиях механизма прерывистой репликации
Обзор
. 2017;93(5):322-338. doi: 10.2183/pjab.93.020. Цунэко Окадзаки
принадлежность
- 1 Нагойский университет.
- PMID: 28496054
- PMCID: PMC5489436
- DOI: 10.2183/пяб.93.020
Обзор
Цунэко Окадзаки. Proc Jpn Acad Ser B Phys Biol Sci.
2017.
Proc Jpn Acad Ser B Phys Biol Sci.
2017.
Автор
Цунэко Окадзаки 1
принадлежность
- 1 Нагойский университет.
- PMID: 28496054
- PMCID: PMC5489436
- DOI: 10.2183/пяб.93.020
Абстрактный
На вилках репликации ДНК общий рост антипараллельных двух дочерних цепей ДНК, по-видимому, происходит в направлении 5′-к-3′ в ведущей цепи и в направлении 3′-к-5′ в отстающей цепи с использованием только ферментной системы, способной удлиняться в направлении от 5′-к-3′, и я описываю в этом обзоре, как мы проанализировали и доказали реакции многоступенчатого синтеза отстающих цепей, называемые механизмом прерывистой репликации, которые включают синтез коротких праймеров РНК, зависимых от праймеров коротких цепей ДНК (Okazaki фрагментов), удаление праймеров из фрагментов Оказаки и заполнение промежутков между фрагментами Окадзаки с помощью РНКазы Н и ДНК-полимеразы I, а также образование длинной отстающей цепи путем соединения фрагментов Оказаки с помощью ДНК-лигазы.
Ключевые слова: ДНК-лигаза; Фрагменты Окадзаки; функция РНКазы Н и ДНК-полимеразы 1; синтез отстающей цепи; праймер-РНК-зависимый синтез фрагментов Оказаки; обработка фрагментов Окадзаки перед лигированием.
Цифры
Рисунок 1.
Модели реакции удлинения…
Рисунок 1.
Модели реакции удлинения дочерних цепей ДНК. (Вариант 1) Одна дочерняя цепь…
Рисунок 1. Модели реакции удлинения дочерних цепей ДНК. (Случай 1) Одна дочерняя цепь синтезируется в направлении от 5′ к 3′, тогда как другая цепь синтезируется в направлении от 3′ к 5′. Обе дочерние цепи синтезируются непрерывно. Эта модель требует нового фермента, который катализирует полимеризацию 3′-к-5′ ДНК. (Случай 2) Обе дочерние цепи синтезируются в направлении от 5′ к 3′ на микроскопическом аналитическом уровне. Лидирующая нить (которая удлиняется в направлении от 5′ к 3′ при наблюдении с помощью макроскопического аналитического метода) синтезируется непрерывно, тогда как отстающая нить (которая, по-видимому, удлиняется в направлении от 3′ к 5′) синтезируется непрерывно. собраны из прерывисто синтезированных небольших фрагментов ДНК.
(Случай 2) Обе дочерние цепи синтезируются в направлении от 5′ к 3′ на микроскопическом аналитическом уровне. Лидирующая нить (которая удлиняется в направлении от 5′ к 3′ при наблюдении с помощью макроскопического аналитического метода) синтезируется непрерывно, тогда как отстающая нить (которая, по-видимому, удлиняется в направлении от 3′ к 5′) синтезируется непрерывно. собраны из прерывисто синтезированных небольших фрагментов ДНК.
Рисунок 2.
Стратегия определения направления…
Рисунок 2.
Стратегия определения направления синтеза ДНК с помощью анализа расщепления экзонуклеазой. Т4…
Фигура 2. Стратегия определения направления синтеза ДНК с помощью анализа расщепления экзонуклеазой. Бактерии, инфицированные фагом Т4, культивируют при 20 ℃ до пика синтеза ДНК.
Рисунок 3.
Фрагменты Окадзаки в Кишечная палочка…
Рисунок 3.
Фрагменты Окадзаки в E. coli . E. coli был помечен импульсами [ 3…
Рисунок 3.Фрагменты Окадзаки в E. coli . E. coli метили импульсным методом [ 3 H]-тимидином при 20 ℃. (а) Импульсная маркировка в течение 10 секунд. (b) Маркировка импульсов в течение 10 секунд с последующей 2-минутной погоней ( , т.е. , в культуральную среду добавляли избыточное количество нерадиоактивного тимидина). (c) Импульсная маркировка в течение 10 секунд с последующей 20-минутной погоней. Затем клеточную ДНК денатурировали и экстрагировали, а фрагменты ДНК разделяли по длине с помощью центрифуги с градиентом 5–20% сахарозы в щелочных условиях. Пик 3 H, наблюдаемый во фракциях 5–7, относится к фрагментам Окадзаки.
Рис. 4.
Фрагменты Окадзаки, наблюдаемые в E. …
…
Рисунок 4.
Фрагменты Окадзаки, обнаруженные в штамме E. coli , инфицированном термочувствительным мутантным штаммом ДНК-лигазы…
Рисунок 4. фрагментов Оказаки наблюдали в E. coli , инфицированных ДНК-лигазой термочувствительного мутантного штамма бактериофага Т4. (а) Эксперимент по мечению импульсов при низкой температуре. Е. coli инфицировали геном 30 (кодирующим ДНК-лигазу) термочувствительного штамма фага Т4 при 20 ℃ в течение 70 минут. Затем бактерии переносили при 30 ℃ и инкубировали в течение 1 минуты с последующим импульсным мечением [ 3 H]-тимидином в течение различных периодов времени. Затем из бактериальных клеток экстрагировали ДНК и анализировали длину фрагментов ДНК с помощью центрифугирования в градиенте сахарозы в щелочных условиях. (b) Эксперимент по мечению импульсов при высокой температуре. Бактерии, инфицированные фагом Т4, были приготовлены в качестве эксперимента с панелью (а), а затем помечены импульсным способом при 43 ℃. Характер оседания ДНК анализировали с помощью центрифугирования в щелочном градиенте сахарозы. (c) Импульсное мечение при высокой температуре с последующей инкубацией при низкой температуре. 9Клетки 0191 E. coli инфицировали геном 30 термочувствительного мутантного штамма фага Т4 и инкубировали при 20 ℃ в течение 70 минут. Затем бактерии переносили при 43 ℃ и инкубировали в течение 2 минут с последующим импульсным мечением [ 3 H]-тимидином в течение одной минуты. После мечения клетки переносили при 30 ℃ и инкубировали в течение различных периодов времени. Затем ДНК экстрагировали и анализировали центрифугированием в щелочном градиенте сахарозы.
Бактерии, инфицированные фагом Т4, были приготовлены в качестве эксперимента с панелью (а), а затем помечены импульсным способом при 43 ℃. Характер оседания ДНК анализировали с помощью центрифугирования в щелочном градиенте сахарозы. (c) Импульсное мечение при высокой температуре с последующей инкубацией при низкой температуре. 9Клетки 0191 E. coli инфицировали геном 30 термочувствительного мутантного штамма фага Т4 и инкубировали при 20 ℃ в течение 70 минут. Затем бактерии переносили при 43 ℃ и инкубировали в течение 2 минут с последующим импульсным мечением [ 3 H]-тимидином в течение одной минуты. После мечения клетки переносили при 30 ℃ и инкубировали в течение различных периодов времени. Затем ДНК экстрагировали и анализировали центрифугированием в щелочном градиенте сахарозы.
Рисунок 5.
Рейджи Оказаки представляет прерывистый…
Рисунок 5.
Рейджи Окадзаки представляет механизм прерывистой репликации на симпозиуме в Колд-Спринг-Харбор…
Рисунок 5.Рейджи Окадзаки представляет механизм прерывистой репликации на симпозиуме в Колд-Спринг-Харбор 19 июня.68.
Рисунок 6.
Определение направления…
Рисунок 6.
Определение направления синтеза фрагмента Оказаки фага Т4 методом экзонуклеазного расщепления…
Рисунок 6. Определение направления синтеза фрагмента фага Т4 Окадзаки с помощью анализа расщепления экзонуклеазой. Бактерии, инфицированные фагом Т4, культивировали при 20°С до пика синтеза ДНК. Затем бактерии охлаждали до 8 ℃ и импульсно метили [ 3 H]-тимидин в течение 6 секунд (для радиоактивной метки растущего конца ДНК) и с [ 14 C]-тимидином в течение 2,5 минут (для радиоактивной метки всей длины фрагментов Окадзаки). Была выделена фракция короткоцепочечной ДНК 9S, обогащенная фрагментами Оказаки, и далее были разделены цепи Уотсона и Крика. Каждую нить подвергали расщеплению экзонуклеазой I E. coli (переваривание 5′-в-3′) и экзонуклеазой Bacillus subtilis (переваривание 5′-в-3′), а 9Радиоактивность 0079 3 H и 14 C, высвобождаемую из ДНК, подсчитывали в различные периоды времени переваривания. Точки данных нанесены на график для процентного высвобождения 3 H и 14 C. Профили высвобождения 3 H показывают, что точка роста расположена на 3′-конце ДНК.
Была выделена фракция короткоцепочечной ДНК 9S, обогащенная фрагментами Оказаки, и далее были разделены цепи Уотсона и Крика. Каждую нить подвергали расщеплению экзонуклеазой I E. coli (переваривание 5′-в-3′) и экзонуклеазой Bacillus subtilis (переваривание 5′-в-3′), а 9Радиоактивность 0079 3 H и 14 C, высвобождаемую из ДНК, подсчитывали в различные периоды времени переваривания. Точки данных нанесены на график для процентного высвобождения 3 H и 14 C. Профили высвобождения 3 H показывают, что точка роста расположена на 3′-конце ДНК.
Рисунок 7.
Короткие фрагменты накапливаются в…
Рисунок 7.
Короткие фрагменты накапливаются в мутантной ДНК-лигазе, а также ДНК…
Рисунок 7. Короткие фрагменты накапливаются в мутантной ДНК-лигазе, а также в мутантах ДНК-полимеразы E.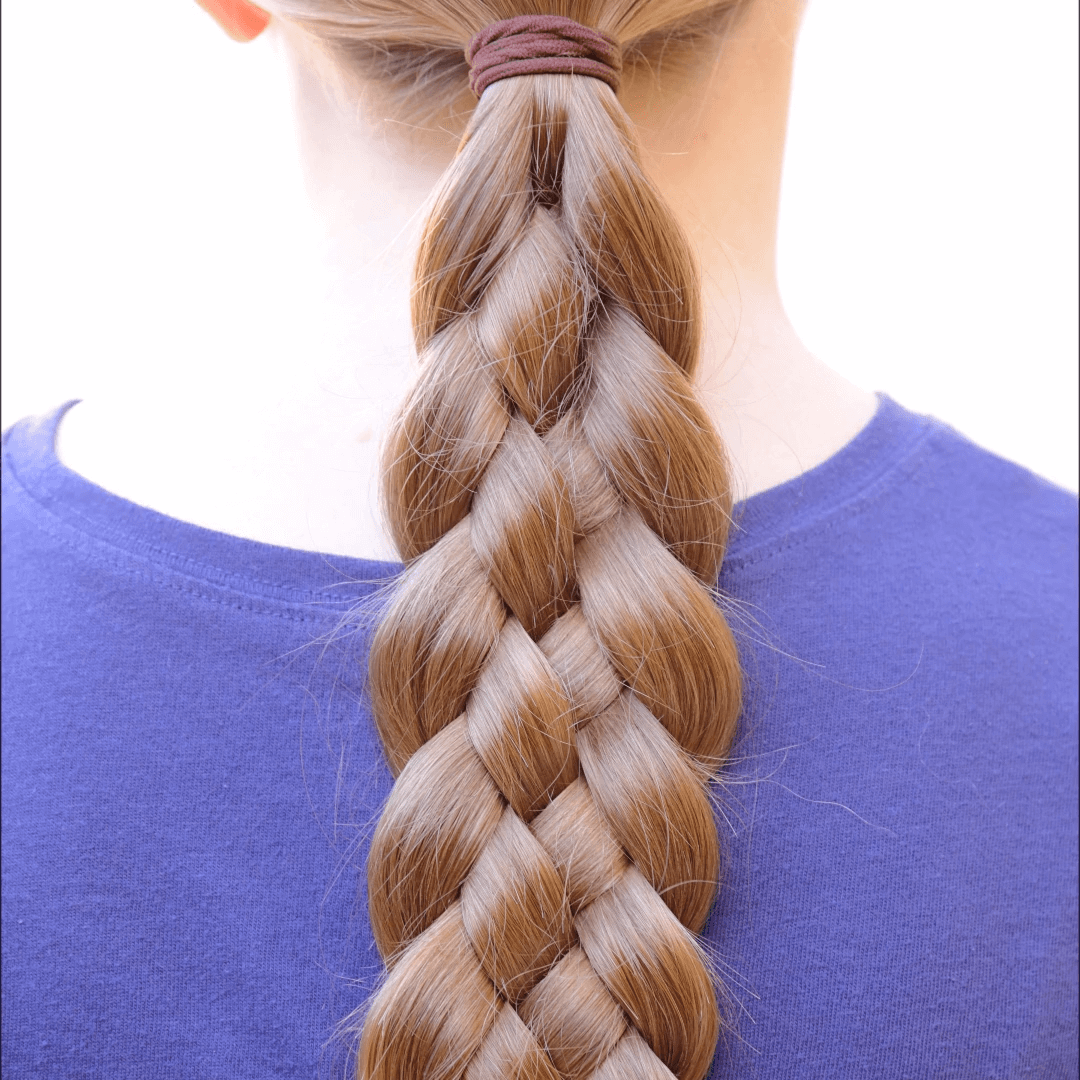 coli в непермиссивном состоянии. Клетки E. coli культивировали при 30 ℃, а затем температуру повышали до 43 ℃, добавляли к [ 3 H]-тимидину на указанные периоды времени для мечения вновь синтезированной ДНК радиоактивным изотопом. Через указанное время ДНК извлекали из клеток и в денатурирующих условиях разделяли по длине с помощью центрифугирования в градиенте 5–20% щелочной сахарозы. Фрагменты Оказаки наблюдались как короткие фрагменты ДНК с коэффициентом седиментации около 10S. Результаты для штамма дикого типа; штамм ligts7, обладающий активностью чувствительной к температуре ДНК-лигазы; polA12 штамм, обладающий термочувствительной полимеразной активностью ДНК-полимеразы I; был показан штамм polAex1 , обладающий чувствительной к температуре экзонуклеазной активностью ДНК-полимеразы I с 5′-на-3′.
coli в непермиссивном состоянии. Клетки E. coli культивировали при 30 ℃, а затем температуру повышали до 43 ℃, добавляли к [ 3 H]-тимидину на указанные периоды времени для мечения вновь синтезированной ДНК радиоактивным изотопом. Через указанное время ДНК извлекали из клеток и в денатурирующих условиях разделяли по длине с помощью центрифугирования в градиенте 5–20% щелочной сахарозы. Фрагменты Оказаки наблюдались как короткие фрагменты ДНК с коэффициентом седиментации около 10S. Результаты для штамма дикого типа; штамм ligts7, обладающий активностью чувствительной к температуре ДНК-лигазы; polA12 штамм, обладающий термочувствительной полимеразной активностью ДНК-полимеразы I; был показан штамм polAex1 , обладающий чувствительной к температуре экзонуклеазной активностью ДНК-полимеразы I с 5′-на-3′.
Рисунок 8.
Скопление осколков Окадзаки в…
Рисунок 8.
Скопление осколков Окадзаки в pol A и rnh мутантный штамм E. coli .…
Рисунок 8. Накопление фрагментов Оказаки в pol A и rnh мутантных штаммов E. coli . Дикий тип, rnh мутант, rnh и polA 5′-3′ экзонуклеазный мутант E. coli выращивали при 30 ℃, а затем переносили на 43 ℃. При этой непермиссивной температуре клетки инкубировали в присутствии [ 3 H]-тимидин в течение указанных периодов времени для радиоактивной метки вновь синтезированной ДНК. После инкубации ДНК экстрагировали в денатурирующих условиях и разделяли по длине центрифугированием в 5–20% щелочном градиенте сахарозы. Фрагменты Оказаки наблюдались как короткие фрагменты ДНК с коэффициентом седиментации около 10S. Вверху слева, штамм дикого типа: вверху справа штамм РНКазы H, обладающий температурно-чувствительной активностью РНКазы H. Внизу слева: двойной мутантный штамм, обладающий чувствительной к температуре 5′-3′-экзонуклеазной активностью ДНК-полимеразы 1 и активностью РНКазы H; внизу справа, polA4113 , который содержал термочувствительную ДНК-полимеразу I, чья экзонуклеазная активность 5′-к-3′ подавлялась при высокой температуре.
Рисунок 9.
Метод выделения грунтовки…
Рисунок 9.
Метод выделения праймерной РНК из фрагментов Оказаки и определение цепи…
Рисунок 9.Метод выделения праймерной РНК из фрагментов Оказаки и определения длины цепи.
Рисунок 10.
Обнаружение интактной РНК-праймера.
Рисунок 10.
Обнаружение интактной РНК-праймера.
Рисунок 10.Обнаружение интактной РНК-праймера.
Рисунок 11.
Ступени прерывистой ДНК…
Рисунок 11.
Этапы реакции прерывистой репликации ДНК. Ведущая нить синтезируется непрерывно…
Рисунок 11.Этапы реакции прерывистой репликации ДНК. Ведущая цепь синтезируется непрерывно, а отстающая цепь синтезируется прерывисто. Реакция элонгации отстающей цепи состоит из пяти стадий: I — раскручивание матрицы ДНК; II, синтез праймеров; III, синтез ДНК (фрагмент Оказаки); IV, деградация праймера и заполнение пробелов; и V, лигирование фрагментов Окадзаки. Точки на матричной ДНК указывают на сигнальные последовательности для синтеза РНК-праймеров.
См. это изображение и информацию об авторских правах в PMC
Похожие статьи
- Комплексы РНК-праймер-примаза служат сигналом для рециклинга полимеразы и инициации фрагмента Оказаки в репликации ДНК фага Т4.
Спиринг М.М., Ханойан П., Ганнаварам С.
 , Бенкович С.Дж.
Спиринг М.М. и др.
Proc Natl Acad Sci U S A. 30 мая 2017 г.; 114 (22): 5635-5640. doi: 10.1073/pnas.1620459114. Epub 2017, 15 мая.
Proc Natl Acad Sci U S A. 2017.
PMID: 28507156
Бесплатная статья ЧВК.
, Бенкович С.Дж.
Спиринг М.М. и др.
Proc Natl Acad Sci U S A. 30 мая 2017 г.; 114 (22): 5635-5640. doi: 10.1073/pnas.1620459114. Epub 2017, 15 мая.
Proc Natl Acad Sci U S A. 2017.
PMID: 28507156
Бесплатная статья ЧВК. - Высвобождение праймера является лимитирующим событием в синтезе отстающей цепи, опосредованном реплисомой Т7.
Эрнандес А.Дж., Ли С.Дж., Ричардсон К.С. Эрнандес А.Дж. и соавт. Proc Natl Acad Sci USA. 2016 May 24;113(21):5916-21. doi: 10.1073/pnas.1604894113. Epub 2016 9 мая. Proc Natl Acad Sci U S A. 2016. PMID: 27162371 Бесплатная статья ЧВК.
- Механизм контроля рециркуляции полимеразы отстающей цепи во время репликации ДНК бактериофага Т4.
Ян Дж., Нельсон С.В., Бенкович С.Дж. Ян Дж.
и др.
Мол Ячейка. 2006 20 января; 21 (2): 153–64. doi: 10.1016/j.molcel.2005.11.029.
Мол Ячейка. 2006.
PMID: 16427006 - Процессы инициации отстающих цепей в репликации ДНК эукариот – цепочки высококоординированных реакций, регулируемые мультибелковыми комплексами.
Nasheuer HP, Onwubiko NO. Нашуер Х.П. и др. Гены (Базель). 2023 29 апреля; 14 (5): 1012. doi: 10.3390/genes14051012. Гены (Базель). 2023. PMID: 37239371 Бесплатная статья ЧВК. Обзор.
- Повторяющийся синтез отстающей цепи ДНК реплисомой бактериофага Т4.
Спиринг М.М., Нельсон С.В., Бенкович С.Дж. Спиринг М.М. и др. Мол Биосист. 2008 ноябрь;4(11):1070-4. дои: 10.1039/b812163j. Epub 2008, 29 сентября. Мол Биосист. 2008. PMID: 18931782 Обзор.
 , Бенкович С.Дж.
Спиринг М.М. и др.
Proc Natl Acad Sci U S A. 30 мая 2017 г.; 114 (22): 5635-5640. doi: 10.1073/pnas.1620459114. Epub 2017, 15 мая.
Proc Natl Acad Sci U S A. 2017.
PMID: 28507156
Бесплатная статья ЧВК.
, Бенкович С.Дж.
Спиринг М.М. и др.
Proc Natl Acad Sci U S A. 30 мая 2017 г.; 114 (22): 5635-5640. doi: 10.1073/pnas.1620459114. Epub 2017, 15 мая.
Proc Natl Acad Sci U S A. 2017.
PMID: 28507156
Бесплатная статья ЧВК. и др.
Мол Ячейка. 2006 20 января; 21 (2): 153–64. doi: 10.1016/j.molcel.2005.11.029.
Мол Ячейка. 2006.
PMID: 16427006
и др.
Мол Ячейка. 2006 20 января; 21 (2): 153–64. doi: 10.1016/j.molcel.2005.11.029.
Мол Ячейка. 2006.
PMID: 16427006
Посмотреть все похожие статьи
Цитируется
- Естественная история ДНК-зависимых ДНК-полимераз: множественные пути к происхождению ДНК.
de Farias ST, Furtado ANM, Dos Santos Junior AP, José MV. де Фариас С.Т. и др. Вирусы. 2023 14 марта; 15 (3): 749. дои: 10.3390/v15030749. Вирусы. 2023. PMID: 36992459 Бесплатная статья ЧВК.
- Везикулы внешней мембраны Dinoroseobacter shibae обогащены сайтом разрешения хромосомных димеров dif .
Ван Х., Бейер Н., Бедекер С., Штайер Х., Хенке П., Нойманн-Шааль М., Мански Дж., Роде М., Оверманн Дж., Петерсен Дж., Клавонн Ф., Куклик М., Энгельманн С., Томаш Дж., Вагнер-Дёблер И. . Ван Х и др. mSystems. 2021 12 января; 6 (1): e00693-20.
doi: 10.1128/mSystems.00693-20.
mSystems. 2021.
PMID: 33436507
Бесплатная статья ЧВК. - Молекулярные механизмы инициации эукариотического происхождения, прогрессии репликационной вилки и поддержания хроматина.
Юань З., Ли Х. Юань Зи и др. Biochem J. 30 сентября 2020 г .; 477 (18): 3499-3525. DOI: 10.1042/BCJ20200065. Биохим Дж. 2020. PMID: 32970141 Бесплатная статья ЧВК.
- Решение 50-летней проблемы фрагмента Окадзаки.
Бургеры ПМ. Бургеры ПМ. Proc Natl Acad Sci USA. 2019 Feb 26;116(9):3358-3360. doi: 10.1073/pnas.1
2116. Epub 2019 15 февраля. Proc Natl Acad Sci U S A. 2019. PMID: 30770440 Бесплатная статья ЧВК. Аннотация недоступна. - Почти непрерывно синтезируемые ведущие цепи в Escherichia coli разрываются путем вырезания рибонуклеотидов.
Кронан Г.Е., Кузьминова Е.А., Кузьминов А. Кронан Г.Э. и соавт. Proc Natl Acad Sci U S A. 2019 Jan 22;116(4):1251-1260. doi: 10.1073/pnas.1814512116. Epub 2019 7 января. Proc Natl Acad Sci U S A. 2019. PMID: 30617079 Бесплатная статья ЧВК.
 doi: 10.1128/mSystems.00693-20.
mSystems. 2021.
PMID: 33436507
Бесплатная статья ЧВК.
doi: 10.1128/mSystems.00693-20.
mSystems. 2021.
PMID: 33436507
Бесплатная статья ЧВК.
Просмотреть все статьи «Цитируется по»
Рекомендации
- Херши А., Чейз М. (1952) Независимые функции вирусного белка и нуклеиновой кислоты в росте бактериофага. J. Gen. Physiol. 36, 39–56. — ЧВК — пабмед
- Уотсон Дж. , Крик Ф. (1953) Молекулярная структура нуклеиновых кислот. Природа 171, 737–738.
—
пабмед
- Уотсон Дж.
- Леман И.Р., Бессман М.Дж., Симмс Э.С., Корнберг А. (1958) Ферментативный синтез дезоксирибонуклеиновой кислоты. 1 Приготовление субстратов и частичная очистка фермента из кишечной палочки. Дж. Биол. хим. 233, 163–170. — пабмед
- Окадзаки Р., Окадзаки Т., Курики Ю. (1960) Выделение рамнозы тимидиндифосфата и нового сахарного соединения тимидиндифосфата из штамма Escherichia coli B.
- Окадзаки Р., Окадзаки Т., Курики Ю. (1960) Выделение рамнозы тимидиндифосфата и нового сахарного соединения тимидиндифосфата из штамма Escherichia coli B.
 , Крик Ф. (1953) Молекулярная структура нуклеиновых кислот. Природа 171, 737–738.
—
пабмед
, Крик Ф. (1953) Молекулярная структура нуклеиновых кислот. Природа 171, 737–738.
—
пабмед