Химическое строение волоса
Химическое строение волос достаточно сложное, как уже упоминалось ранее волос состоит из белка, который растет из клеток внутри волосяного фолликула. Как только образуются живые клетки внутри волоса, они начинают свой путь вверх через волосяной фолликул. Вновь образованные клетки волоса созревают и при этом наполняются волокнистым белком, который носит название кератин. Данный процесс созревания — ороговение или кератинизация. После того, как клетки наполняются кератином, они продвигаются вверх, при этом теряют ядро и умирают. К тому времени как волос выходит наружу из фолликула клетки волоса уже полностью ороговели и не являются живыми.
Волосы состоят на 90% из белка — кератина. Белок состоит из длинных цепей аминокислот, которые в свою очередь состоят из химических элементов. Основными химическими элементами, из которых состоит человеческий волос являются: углерод, кислород, водород, азот и сера. Из этих элементов так же состоят кожа и ногти.
Процентное содержание в нормальном волосе
Углерод — 51%
Кислород — 21%
Водород — 6%
Азот — 17%
Сера — 5%
Цепочка кератина
Белки состоят из длинных цепочек аминокислот, которые соединились друг с другом как цепочки бисера. Сильная химическая связь, которая соединяет аминокислоты между собой носит название — белковой или полипептидной. Белки — это длинные, спиральные сложные аминокислотные цепочки, соединенные белковыми связями. Белковые цепочки кератина переплетаются друг с другом и образуют гибкую спираль. Именно такая сложная структура кератина делает его прочным.
Кортекс волоса состоит из миллионов сложных белковых цепочек. Эти цепи сшиты как ступеньки на лестнице несколькими типами дополнительных боковых связей. Именно они связывают цепочки кератина вместе и отвечают за сильную прочность и хорошую эластичность человеческого волоса.
Химические связи в волосах
Водородная связь является очень слабой физической поперечной связью, которая легко разрушается водой и температурой. И хотя по отдельности водородные связи очень слабые, но их в волосах так много, что на их долю приходиться около одной трети от общей численности связей в волосах. Водородные связи легко разрушаются при намокании волос. Это позволяет волосам легко растягиваться. При высыхании волоса водородные связи восстанавливаются и фиксируют форму волос в том положении в котором волосы оказались.
Ионные или солевые связи так же являются слабыми физическими связями, образующимися поперек волоса между соседними белковыми цепочками. Ионные связи зависят от pH на волосах, поэтому они легко разрушаются сильными щелочными и кислыми растворами.
Дисульфидная связь — это сильная химическая связь, которая очень отличается от физических водородной и ионной. Она соединяет атомы серы двух соседних аминокислот цистеина, что бы создать одну — цистин. Цистин объединяет две белковые цепочки в единое целое.
Хотя самих дисульфидных связей гораздо меньше, чем водородных и ионных, но они очень сильные и не разрушаются водой. Разорвать дисульфидную связь можно с помощью препаратов для химической завивки и выпрямления. Мало того нормальное температурное воздействие, как, например, при термоукладке, не разрушают дисульфидных связей. Однако очень сильные воздействия температуры, как при воздействии кипятка и некоторых высокотемпературных инструментов для укладки, нарушают эти связи. Яркий пример такого сильного воздействия — утюжки для выпрямления волос.
Большинство препаратов для химической завивке и выпрямлении на украинском рынке вызывают разрушение дисульфидные связи с возможностью дальнейшего восстановления с помощью нейтрализаторов или фиксажа. Однако, все еще на зарубежных рынках, таких как Европа, Америка и т. п. Можно встретить специальные препараты для химического выпрямления, которые разламывают дисульфидные связи без возможности их дальнейшего восстановления.
Однако, все еще на зарубежных рынках, таких как Европа, Америка и т. п. Можно встретить специальные препараты для химического выпрямления, которые разламывают дисульфидные связи без возможности их дальнейшего восстановления.
фазы роста, жизненный цикл, химический состав, виды связей, пигмент
- Изменение структуры волос на протяжении жизни человека
- Формирование фолликул и начало роста волос
- Фазы роста волос
- Форма волос
- Строение волоса
- Химический состав
- Пигмент
 org/ListItem»> Виды связей
org/ListItem»> Виды связейВолосы растут на всём кожном покрове человека, за исключением ладоней и ступней. На теле нашем есть участки с густой растительностью, а также еле заметными почти прозрачными волосками. Изначально, их роль заключалась в защите. Но в процессе эволюции функции изменились и стали атрибутом декоративности, нежели необходимости.
Чтобы эффективно выполнять различные парикмахерские процедуры, такие как окрашивание, обесцвечивание, химическая завивка или выпрямление, нужно знать, из чего состоит волос, какие элементы участвуют в реакции при нанесении красителя, ухаживающих или меняющих форму веществ. Давайте узнаем о строении волоса человека, как и из чего формируется его колба, фолликул, форма и цвет.
Изменение структуры волос на протяжении жизни человека
Во время наблюдений учёными за процессом роста волос человека, было установлено, что их формирование происходит ещё до рождения, а на разных этапах жизни их структура меняется. Как это происходит?
Как это происходит?
- Сначала появляются пушковые. Они вырастают при рождении и остаются у ребёнка до возраста трёх лет. Название само говорит, что их структура очень мягкая и тонкая.
- В возрасте от трёх до шестнадцати лет у ребёнка начинают формироваться подростковые волосы, которые развиваются в генетически заложенный тип и уже можно определить, какими они будут в дальнейшем.
- В юношеском возрасте — 16–17 лет, волосы переходят в стадию зрелости. А к 18 годам, формирование их структуры окончательно завершается.
В период полового созревания у мужчин появляется растительность на теле и лице — усы, борода. А у женщин — только на теле. К старости у пожилых людей наблюдается интенсивный рост бровей, появляются волосы в ушах и увеличивается их количество в носу. С возрастом структура становится жёстче, а луковица углубляется в кожу.
Однако, то, что волосы продолжают расти и после смерти — это миф. Такое предположение изначально сделали наблюдатели за эксгумацией, когда видели их неизменными после раскрытия могил. Но врачи-патологоанатомы опровергли такую возможность, объяснив это явление тем, что ткань волоса разлагается гораздо дольше остальных органов и не подвергается воздействию паразитов.
Но врачи-патологоанатомы опровергли такую возможность, объяснив это явление тем, что ткань волоса разлагается гораздо дольше остальных органов и не подвергается воздействию паразитов.
Формирование фолликул и начало роста волос
Рост начинается из-за активного деления клеток в наружном слое кожи. Такое их разрастание называется «зачаток волоса», который затем принимает каплевидную форму и впоследствии становится фолликулом. Когда капля (язычок) углубляется в кожу, клетка продолжает делиться. Это разросшееся накопление клеток превращается в волосяной сосочек, который обрастает белком и получается головка с заострённым кончиком — луковица. Таким образом, белок в составе волос образуется ещё до появления его на поверхности кожи. В строении фолликула, а затем и луковицы участвуют клетки — меланоциты, в которых закладывается будущий пигмент, а также белок — кератин, состоящий из аминокислот.
Питание корня и приток строительных веществ осуществляется через соединённый с кровеносным сосудом сосочек, из которого и начинается рост волоса.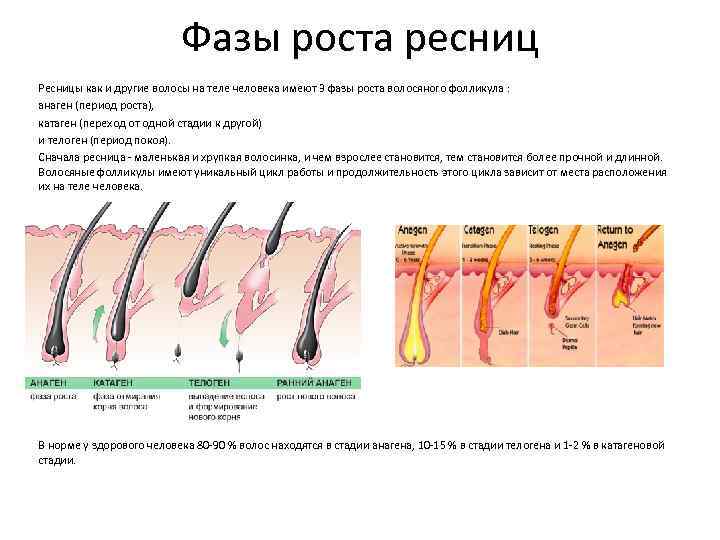 Фолликул роговеет и через образовавшееся отверстие в его сердцевине появляется маленький отросток, который пробившись сквозь слой кожи, выбирается наружу.
Фолликул роговеет и через образовавшееся отверстие в его сердцевине появляется маленький отросток, который пробившись сквозь слой кожи, выбирается наружу.
Новый (первичный) волос называется сосочковым, поскольку питается напрямую из кровеносного сосуда. Постепенно подача кератина из сосочка прекращается, волос врастает в канал сильнее, его рост останавливается и теряет связь с питающим органом, а луковица роговеет окончательно. В этом состоянии волосы называются колбовидными. Они долго остаются в коже, поэтому большая часть растительности человека состоит именно из них.
Затем, клеточная масса, расположенная под колбовидными волосами, начинает опять делиться и образовывает новую ткань, которая выталкивает старую. Выросший на месте выпавшего молодой волос называется вторичным. Это непрерывный естественный процесс. Ежедневная норма выпадения составляет 60–100 волос. Максимальная продолжительность их жизни — 6 лет.
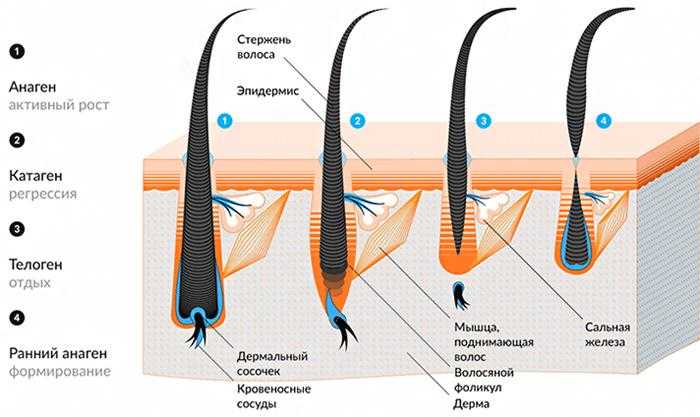
Фазы роста волос
Поскольку замена старых волос на новые — процесс постоянный, то у него есть определённый цикл, который состоит из нескольких фаз:
- анаген;
- катаген;
- телоген;
- ранний анаген.


Что они означают?
- Анаген — фаза активного деления клеток в волосяном мешочке. В период роста луковицы в коже вокруг неё начинается синтез рибонуклеиновой кислоты. В этой фазе волосяной сосочек получает наиболее интенсивное питание из кровеносных сосудов. В то же время происходит соединение луковицы с нервными окончаниями — поэтому и возникает боль при вырывании волоска с корнем. В фазе анагена также закладывается пигмент. Эта стадия роста полностью формирует луковицу и будущий волос, период жизни которого будет длиться от двух до шести лет.
- Катаген — промежуточная фаза, которая наступает после активного роста. В этот период прекращается подача питательных веществ к сосочку и синтез меланоцитов. Рост волос останавливается и происходит ороговение фолликул. Промежуточная стадия длится 2–3 недели.
- Телоген — фаза покоя, когда луковица становится полностью неактивной. Питание заканчивается и сосочек переходит в состояние покоя, но волосяная колба находится на поверхности ещё 2–3 месяца, а затем безболезненно покидает свой мешочек. Ежедневно выпадает около ста телогеновых волос. Общая длительность этой фазы может составлять до полугода.
- Ранний анаген — это период активного формирования вторичного волоса под ещё не выпавшей старой колбой.
 Ежедневно выпадает около ста телогеновых волос. Общая длительность этой фазы может составлять до полугода.
Ежедневно выпадает около ста телогеновых волос. Общая длительность этой фазы может составлять до полугода.Этот процесс непрерывен, но с возрастом он замедляется, а фаза анагена укорачивается. Со временем часть фолликулов теряют способность к формированию новых волос. А в некоторых случаях их большинство совсем отмирает и наступает алопеция — частичное, а затем и полное облысение.
Форма волос
То, каким будет волос — прямым, волнистым или сильно кучерявым зависит от формы канала, в котором зарождается луковица.
- Ровный волос формируется из прямого канала.
- Волнистый — из серповидного. Чем сильнее загнут канал, тем больше кудрявость.
- Тугие негроидные кудряшки вырастают из спиральных каналов.
Таким образом, будущая форма волоса закладывается ещё до того, как он появится на поверхности кожи.
Если посмотреть под микроскопом на волосы разных типов в поперечном сечении, то можно заметить следующие различия.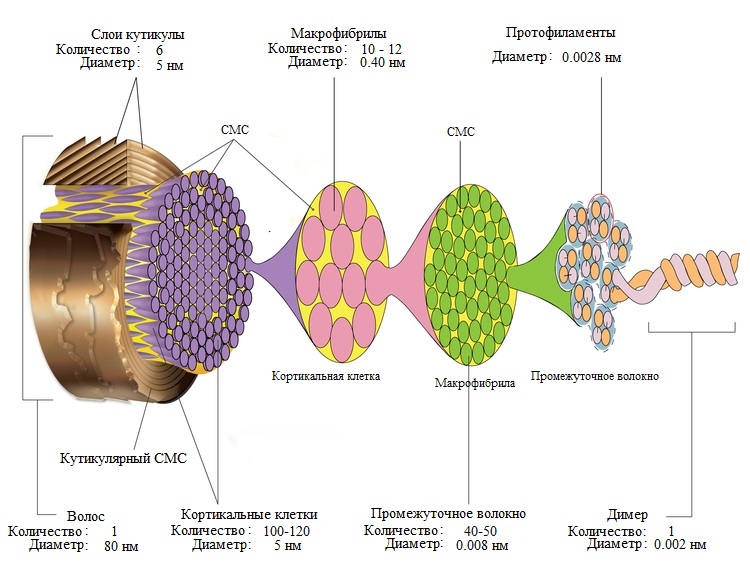
- Азиатские чаще всего толстые, прямые и жёсткие, а их поперечное сечение — круглое.
- Европейские обычно средней толщины, слегка волнистые, сечение — эллипсообразное.
- Негроидные — жёсткие, кучерявые, в разрезе имеют лентообразную форму.
Форма канала влияет не только на то, каким будет волос, но и на его поперечное сечение.
Строение волоса
При рассмотрении разреза волоса человека через микроскоп, можно увидеть, что он состоит из трёх основных слоёв:
- чешуи — кутикулы;
- коры — кортекса;
- и сердцевины — стержня, мозгового слоя или медулы.
А также имеет цилиндрическую форму.
Рассмотрим подробнее строение всех слоёв.
- Кутикула волоса (чешуя) — это защитный слой от влияния извне. Она состоит из роговых листиков, расположенных в шахматном порядке, которые похожи на еловую шишку или рыбью чешую. Эти лепестки чувствительны к щелочному или кислотному воздействию.
- У кортекса строение сложное. Он формируется из удлинённых волокон, переплетающихся между собой. В этом слое находится пигмент, водородные и серные связи, а также питательные вещества.
- Строение стержня (медулы) образуется из мозговых клеток, составляющих основу колбы. К этой сердцевине крепятся остальные слои.
 Он формируется из удлинённых волокон, переплетающихся между собой. В этом слое находится пигмент, водородные и серные связи, а также питательные вещества.
Он формируется из удлинённых волокон, переплетающихся между собой. В этом слое находится пигмент, водородные и серные связи, а также питательные вещества.Помимо этого, в формировании волоса участвует также жировая железа и мышца.
- Жировая железа отвечает за эластичность, предотвращение ороговения волос и кожи головы. А также выработка сального секрета — естественной жировой плёнки, защищает от проникновения внутрь поверхностных загрязнений.
- Мышца прижимает волосы к коже головы, создавая защиту от холода, или при нервном потрясении. Образное выражение — «волосы встали дыбом», становится реальным благодаря именно этой мышце.
Каждая составная часть в строении волоса отвечает за отдельную функцию.
Строение волоса человека
Химический состав
Человек весь состоит из элементов периодической таблицы Менделеева. Химический состав волос нужно знать, чтобы предугадывать реакцию различных препаратов при выполнении процедур долговременной укладки и окрашивания.
Химический состав волос нужно знать, чтобы предугадывать реакцию различных препаратов при выполнении процедур долговременной укладки и окрашивания.
Соотношение химических элементов в волосе, следующее:
- углерод (C) — 49,6%;
- кислород (O) — 23,2%;
- азот (N) — 16,8%;
- водород (H) — 6,4%;
- сера (S) — 4%.
Они влияют за качество окрашивания, стойкость пигмента, а также формирование или выпрямление завитка.
Вещества, из которых слагается волос, образуют фибриллярный белок — кератин и все его аминокислоты. Это основной строительный материал.
Кератин состоит из следующих аминокислот:
- аланин;
- аргинин;
- аспарагиновая кислота;
- валин;
- глицин;
- гистидин;
- глютаминовая кислота;
- изолейцин;
- лейцин;
- лизин;
- метионин;
- пролин;
- серин;
- тирозин;
- треонин;
- триптофан;
- фенилаланин;
- цистин.
Недостаток хотя бы одной аминокислоты в человеческом волосе ведёт к их ломкости, а луковицы — к необратимому отмиранию и выпадению. Кератин обладает высокой механической прочностью, уступая лишь хитину, который образует ороговевшие части животных.
Кератин обладает высокой механической прочностью, уступая лишь хитину, который образует ороговевшие части животных.
Виды связей
При смене цвета в результате окрашивания, или формы волоса при завивке или выпрямлении, меняется его водородная или серная связь. Что это такое и какое влияние на неё оказывают химические вещества?
- Водородные (продольные) связи — это одинарные мостики, расположенные вдоль волоса. Они отвечают за изменение натурального или косметического пигмента. Реакция красителей основывается на воздействии оксида водорода с волокнистым веществом кортекса.
- Серные (поперечные) связи — это двойные мостики, расположенные поперёк волоса. Они менее хрупкие, чем водородные. Отвечают за форму — чем сильнее дисульфидная (двойная серная) связь, тем завиток интенсивнее. Основная реакция препаратов для долговременной укладки заложена в воздействии серосодержащих составов с этими мостиками в кортексе.
Все современные химические средства синтезированы из компонентов, наиболее сходных по составу с человеческим волосом, чтобы они встраивались в его волокна и не разрывали связи, а деликатно их растягивали.
Пигмент
Это цвет, который видит глаз человека. Он закладывается ещё до рождения и зависит от генетических особенностей. Цвет образуется в кортексе из клеток меланоцитов, которые представляют два вида натуральных пигментов:
- меланин — чёрный или коричневый;
- феомеланин — красный или жёлтый.
Натуральная база образуется при сочетании данных типов пигментов. Это цвет, который даётся человеку при рождении.
Но пигменты могут быть и косметическими — это приобретённый цвет при помощи окрашивания. При создании косметической базы, молекулы красителя выталкивают естественный пигмент и встраиваются в образовавшиеся пустоты.
Подытожим, напомнив основные тезисы. Волосы покрывают почти всё тело, за исключением стоп и ладоней. Их структура и строение зависят от периода жизни человека. У новорождённых детей они пушковые, а после трёх лет превращаются в подростковые. Окончательное формирование завершается к половому созреванию. Зарождение волоса происходит ещё до рождения ребёнка. Его цвет и форма зависят от генетической расположенности человека. Прежде чем волосяной кончик появится на поверхности, внутри кожного покрова происходят сложные биохимические процессы деления клеток, формирования из них сосочка, соединённого с кровеносным сосудом, а затем обрастающего нервными окончаниями. Рост и выпадение волос — процесс естественный и непрерывный. Цикл состоит из трёх основных фаз — анаген, катаген, телоген, а также одной добавочной — ранний анаген. Суточная норма выпадения у взрослого человека — 60–100 штук. Таким образом, рост новых волос не прекращается до конца жизни.
Его цвет и форма зависят от генетической расположенности человека. Прежде чем волосяной кончик появится на поверхности, внутри кожного покрова происходят сложные биохимические процессы деления клеток, формирования из них сосочка, соединённого с кровеносным сосудом, а затем обрастающего нервными окончаниями. Рост и выпадение волос — процесс естественный и непрерывный. Цикл состоит из трёх основных фаз — анаген, катаген, телоген, а также одной добавочной — ранний анаген. Суточная норма выпадения у взрослого человека — 60–100 штук. Таким образом, рост новых волос не прекращается до конца жизни.
Строение волос — кутикула
Строение волос — кортекс и медула
Химические свойства волос. В основном состоит из кератина, волосы… | by Beauty Tomorrow
Волосы, в основном состоящие из кератина, также содержат другие элементы и молекулы, влияющие на их внешний вид и поведение
Общий химический состав волос: 45 % углерода, 28 % кислорода, 15 % азота, 7 % водорода и 5 % серы.
Стержень волоса в основном состоит из кератина. Кератин волос жесткий, компактный и сильный. Этот волокнистый белок постепенно образуется внутри клеток из зародышевого слоя. Эти клетки кутикулы характеризуются наличием аморфного кератина, в то время как клетки коры имеют структуру нитей, окруженных кератиновым веществом, более богатым серой и содержащим аминокислоты. Кератин этих нитей образует спираль с расстоянием между витками 0,51 нанометра и структурой, поддерживаемой водородными связями. Этот белок играет ключевую роль в сплоченности и физических свойствах волос.
Волосы в основном состоят из кератина, молекулы со спиральной структурой. © L’Oréal Research Волосы также содержат воду (от 12 до 15 %) и следы минеральных элементов (кальций, кадмий, хром, медь, цинк, железо и кремний). Эти элементы могут быть доставлены к основанию волосяного фолликула посредством кровообращения, а затем способствовать построению стержня волоса. Помимо этого вклада, окружающая среда может, например, из-за загрязнения, быть источником определенных элементов, таких как свинец.
Волосы также содержат липидные компоненты (3% от их состава). Они производятся в волосяной луковице из стеролов, жирных кислот и керамидов. Присутствуя в основном в межклеточном цементе кортекса и кутикулы, они придают волосам определенную непроницаемость и обеспечивают сцепление волосяного волокна.
Другие липиды получают из секрета сальных желез: кожного сала. Кожное сало образуется из зрелых сальных клеток, которые лопнули, и в основном содержит липиды (триглицериды, воск, сквален, этерифицированный холестерин и свободный холестерин). Наиболее распространенные триглицериды подвергаются частичному гидролизу бактериями, населяющими кожу головы, Propionibacterium acnes и Propionibacterium granulosum . Этот гидролиз высвобождает свободные жирные кислоты, ди- и моноглицериды и глицерин. Жирные кислоты характеризуются четными и неравномерными по количеству (от С11 до С19) углеродными цепями с многочисленными ненасыщенными участками и разветвлениями. Воски представляют собой сложные эфиры кислот и длинноцепочечных спиртов. Воски представляют собой неполярные соединения, которые метаболизируются флорой и мало подвержены окислению.
Воски представляют собой сложные эфиры кислот и длинноцепочечных спиртов. Воски представляют собой неполярные соединения, которые метаболизируются флорой и мало подвержены окислению.
В сальных железах молекулы ацетата конденсируются с образованием мевалоновой кислоты, изомера изопреноида, и сквалена, линейного углеводорода, состоящего из 30 атомов углерода. Стерины этерифицируются жирными кислотами. Соотношение сквален/холестерин в поверхностной гидролипидной пленке является зеркалом биологической активности сальных желез и эпидермиса. Липидная смесь, которая образует эту пленку на поверхности кожи, смазывает волосы и, таким образом, сохраняет их эластичность и блеск. Из-за зависимости от гормонов кожное сало может вырабатываться в избытке, и волосы становятся жирными и тяжелыми. С другой стороны, если секрета слишком мало, волосы становятся сухими, тусклыми и поврежденными.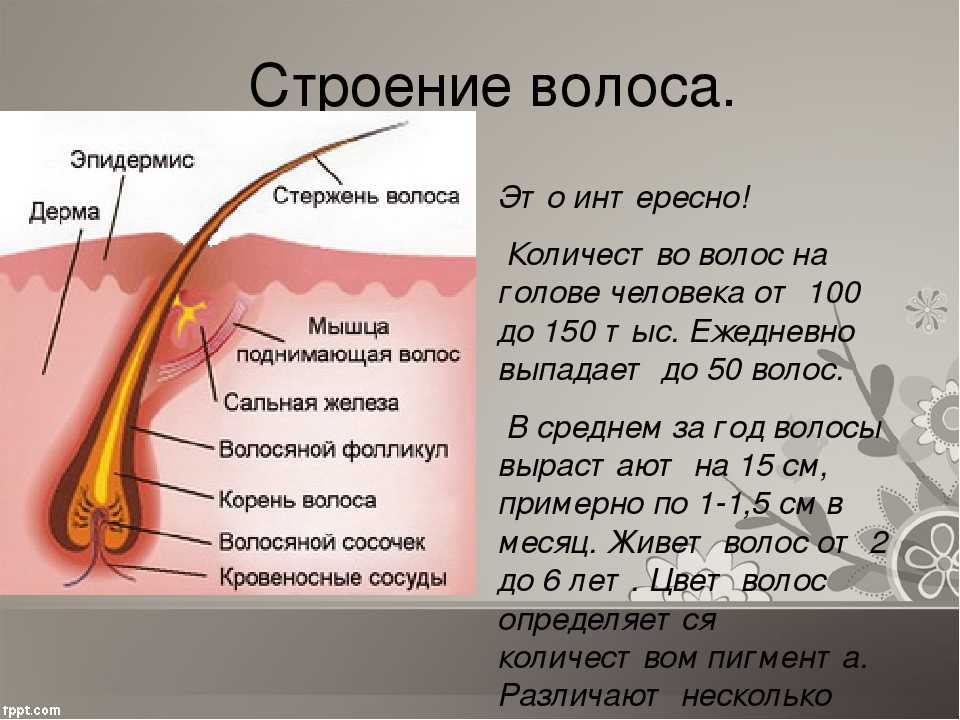
Структура волос человека
Armstrong et al. (2013) Армстронг С.Л., Марквардт Д., Дайс Х., Кучерка Н., Ямани З., Харрун Т.А., Катсарас Дж., Ши А.С., Райнштедтер М.С. Наблюдение высокоупорядоченных доменов в мембранах с холестерином. ПЛОС ОДИН. 2013;8:e619. doi: 10.1371/journal.pone.0066162. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Astbury & Sisson (1935) Astbury WT, Sisson WA. Рентгенологическое исследование структуры волос, шерсти и связанных с ними волокон. III. Конфигурация молекулы кератина и ее ориентация в биологической клетке. Труды Лондонского королевского общества. Серия А, Математические и физические науки. 1935;150:533–551. doi: 10.1098/rspa.1935.0121. [CrossRef] [Google Scholar]
Astbury & Street (1932) Astbury WT, Street A. Рентгеновские исследования структуры волос, шерсти и связанных с ними волокон. I. Генерал. Философские труды Лондонского королевского общества. Серия A, содержащая документы математического или физического характера.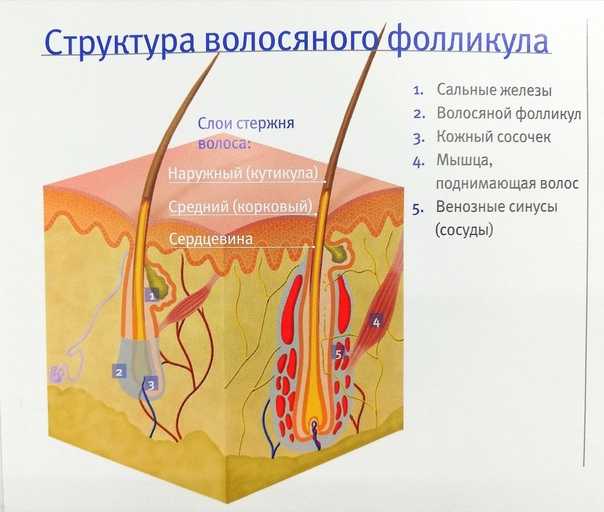 1932; 230: 75–101. doi: 10.1098/rsta.1932.0003. [CrossRef] [Google Scholar]
1932; 230: 75–101. doi: 10.1098/rsta.1932.0003. [CrossRef] [Google Scholar]
Astbury & Woods (1934) Astbury WT, Woods HJ. Рентгенологическое исследование структуры волос, шерсти и связанных с ними волокон. II. Молекулярная структура и эластичные свойства кератина волос. Философские труды Лондонского королевского общества. Серия A, содержащая документы математического или физического характера. 1934;232:333–394. doi: 10.1098/rsta.1934.0010. [CrossRef] [Google Scholar]
Barrett et al. (2012) Барретт М.А., Чжэн С., Рошанкар Г., Олсоп Р.Дж., Белангер Р.К.Р., Хюинь С., Кучерка Н., Райнштадтер М.С. Взаимодействие аспирина (ацетилсалициловой кислоты) с липидными мембранами. ПЛОС ОДИН. 2012;7:e619. doi: 10.1371/journal.pone.0034357. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Barrett et al. (2013) Барретт М.А., Чжэн С., Топпозини Л.А., Олсоп Р.Дж., Дайс Х., Ван А., Яго Н., Мур М., Райнштедтер М.С. Растворимость холестерина в липидных мембранах и образование несмешивающихся холестериновых бляшек при высоких концентрациях холестерина.
Berg, Rosenberg & Asbury (1972) Berg BO, Rosenberg SH, Asbury AK. Гигантская аксональная невропатия. Педиатрия. 1972; 49: 894–899. [PubMed] [Google Scholar]
Briki et al. (2000) Briki F, Busson B, Kreplak L, Dumas P, Doucet J. Исследование биологической ткани от атомного до макроскопического масштаба с использованием синхротронного излучения: пример волос. Клеточная и молекулярная биология. 2000;46:1005–1016. [PubMed] [Академия Google]
Брики и др. (1999) Briki F, Busson B, Salicru B, Estève F, Doucet J. Диагностика рака молочной железы с помощью волос. Природа. 1999; 400: 226–226. дои: 10.1038/22244. [PubMed] [CrossRef] [Google Scholar]
Busson, Engstrom & Doucet (1999) Busson B, Engstrom P, Doucet J. Существование различных структурных зон в ороговевших тканях, выявленное с помощью рентгеновской микродифракции. Журнал синхротронного излучения. 1999; 6: 1021–1030. doi: 10.
Cohen & Parry (1994) Коэн С., Парри Д.А. Альфа-спиральные катушки: больше фактов и лучшие прогнозы. Наука. 1994; 263:488–489. doi: 10.1126/science.8290957. [PubMed] [CrossRef] [Google Scholar]
Creasy et al. (2013) Creasy RK, Resnik R, Iams JD, Lockwood CJ, Greene MF, редакторы. Медицина матери и плода Кризи и Резника: принципы и практика. В. Б. Сондерс; 2013. [Google Scholar]
Crewther et al. (1983) Crewther WG, Dowling LM, Steinert PM, Parry DAD. Структура промежуточных филаментов. Международный журнал биологических макромолекул. 1983;5:267–274. doi: 10.1016/0141-8130(83)
-5. [CrossRef] [Google Scholar]
Crick (1952) Crick FHC. Является ли α -кератин спиральной спиралью? Природа. 1952; 170: 882–883. дои: 10.1038/170882b0. [PubMed] [CrossRef] [Google Scholar]
Franbourg et al. (2003) Franbourg A, Hallegot P, Baltenneck F, Toutaina C, Leroy F. Текущие исследования этнических волос. Журнал Американской академии дерматологии. 2003;48:S115–S119. doi: 10.1067/mjd.2003.277. [PubMed] [CrossRef] [Академия Google]
Фрейзер и др. (1986) Fraser RD, MacRae TP, Parry DA, Suzuki E. Промежуточные филаменты в альфа-кератинах. Труды Национальной академии наук Соединенных Штатов Америки. 1986; 83: 1179–1183. doi: 10.1073/pnas.83.5.1179. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Fraser, MacRae & Rogers (1962) Fraser RD, MacRae TP, Rogers GE. Молекулярная организация альфа-кератина. Природа. 1962; 193: 1052–1055. дои: 10.1038/1931052a0. [PubMed] [CrossRef] [Академия Google]
Фрейзер и др. (1988) Fraser RDB, MacRae TP, Sparrow LG, Parry DAD. Дисульфидное связывание в α -кератине. Международный журнал биологических макромолекул. 1988; 10: 106–112. doi: 10.1016/0141-8130(88)-7. [CrossRef] [Google Scholar]
Furt, Simon-Plas & Mongrand (2011) Furt F, Simon-Plas F, Mongrand S. In: Плазматическая мембрана растений. Мерфи А.С., Шульц Б., Пир В., редакторы. об. 19. Берлин, Гейдельберг: Springer-Verlag; 2011. С. 57–85. (Монографии о растительных клетках). [Перекрестная ссылка] [Академия Google]
С. 57–85. (Монографии о растительных клетках). [Перекрестная ссылка] [Академия Google]
Харроун и др. (1999) Harroun TA, Heller WT, Weiss TM, Yang L, Huang HW. Экспериментальные доказательства гидрофобного соответствия и мембранно-опосредованных взаимодействий в липидных бислоях, содержащих грамицидин. Биофизический журнал. 1999; 76: 937–945. doi: 10.1016/S0006-3495(99)77257-7. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Howell et al. (2000) Хауэлл А., Гроссманн Дж. Г., Чунг К. С., Канби Л., Д. Гарет Р. Е., Хаснайн С. С. Можно ли использовать волосы для скрининга рака молочной железы? Журнал медицинской генетики. 2000;37:297–298. doi: 10.1136/jmg.37.4.297. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Iida & Noma (1993) Iida A, Noma T. Синхротронный рентгеновский музонд и его применение для анализа человеческих волос. Ядерные приборы и методы в физических исследованиях Раздел B: Взаимодействие пучков с материалами и атомами. 1993; 82: 129–138. doi: 10.1016/0168-583X(93)95092-J. [CrossRef] [Google Scholar]
doi: 10.1016/0168-583X(93)95092-J. [CrossRef] [Google Scholar]
Джеймс (2001) Джеймс В. Важность хороших изображений при использовании волос для скрининга рака молочной железы. Журнал медицинской генетики. 2001;38:e619. doi: 10.1136/jmg.38.5.e16. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Veronica & Amemiya (1998) James VJ, Amemiya Y. Упаковка промежуточных нитей в α -кератине пера ехидны. Журнал текстильных исследований. 1998; 68: 167–170. doi: 10.1177/004051759806800303. [CrossRef] [Google Scholar]
Джеймс и др. (1999) Джеймс В., Кирсли Дж., Ирвинг Т., Амемия Й., Куксон Д. Использование волос для скрининга рака молочной железы. Природа. 1999; 398:33–34. дои: 10.1038/17949. [PubMed] [CrossRef] [Академия Google]
Каджиура и др. (2006) Kajiura Y, Watanabe S, Itou T, Nakamura K, Iida A, Inoue K, Yagi N, Shinohara Y, Amemiya Y. Структурный анализ одиночных волокон человеческого волоса путем сканирования микролучевых саксофонов. Журнал структурной биологии. 2006; 155: 438–444. doi: 10.1016/j.jsb.2006.04.008. [PubMed] [CrossRef] [Google Scholar]
Журнал структурной биологии. 2006; 155: 438–444. doi: 10.1016/j.jsb.2006.04.008. [PubMed] [CrossRef] [Google Scholar]
Katsaras et al. (1995) Katsaras J, Raghunathan VA, Dufourc EJ, Dufourcq J. Доказательства двумерной молекулярной решетки в бислоях dppc субгелевой фазы. Биохимия. 1995;34:4684–4688. doi: 10.1021/bi00014a023. [PubMed] [CrossRef] [Google Scholar]
Kreplak et al. (2001a) Kreplak L, Briki F, Duvault Y, Doucet J, Merigoux C, Leroy F, Lévêque JL, Miller L, Carr GL, Williams GP, Dumas P. Профилирование липидов в поперечных стрижках кавказских и афроамериканских волос с использованием синхротрона инфракрасная микроспектрометрия. Международный журнал косметической науки. 2001а; 23:369–374. doi: 10.1046/j.0412-5463.2001.00118.x. [PubMed] [CrossRef] [Google Scholar]
Kreplak et al. (2001b) Креплак Л., Меригу С., Брики Ф., Флот Д., Дусе Дж. Исследование структуры кутикулы человеческого волоса с помощью микродифракции: прямое наблюдение за набуханием комплекса клеточной мембраны. Biochimica et Biophysica Acta (BBA) — структура белка и молекулярная энзимология. 2001b; 1547: 268–274. дои: 10.1016/S0167-4838(01)00195-9. [PubMed] [CrossRef] [Google Scholar]
Biochimica et Biophysica Acta (BBA) — структура белка и молекулярная энзимология. 2001b; 1547: 268–274. дои: 10.1016/S0167-4838(01)00195-9. [PubMed] [CrossRef] [Google Scholar]
Kučerka et al. (2005) Кучерка Н., Лю Ю., Чу Н., Петраче Х.И., Тристрам-Нэгл С., Нэгл Дж. Ф. Структура липидных бислоев полностью гидратированной жидкой фазы DMPC и DLPC с использованием рассеяния рентгеновских лучей от ориентированных многослойных массивов и от однослойных везикул. Биофизический журнал. 2005; 88: 2626–2637. doi: 10.1529/biophysj.104.056606. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Kuč, Tristram-Nagle & Nagle (2006) Kučerka N, Tristram-Nagle S, Nagle JF. Более пристальный взгляд на структуру полностью гидратированных двойных слоев dppc жидкой фазы. Биофизический журнал. 2006;90:L83–L85. doi: 10.1529/biophysj.106.086017. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Lupas & Gruber (2005) Lupas AN, Gruber M. Структура α -спиральных спиральных катушек.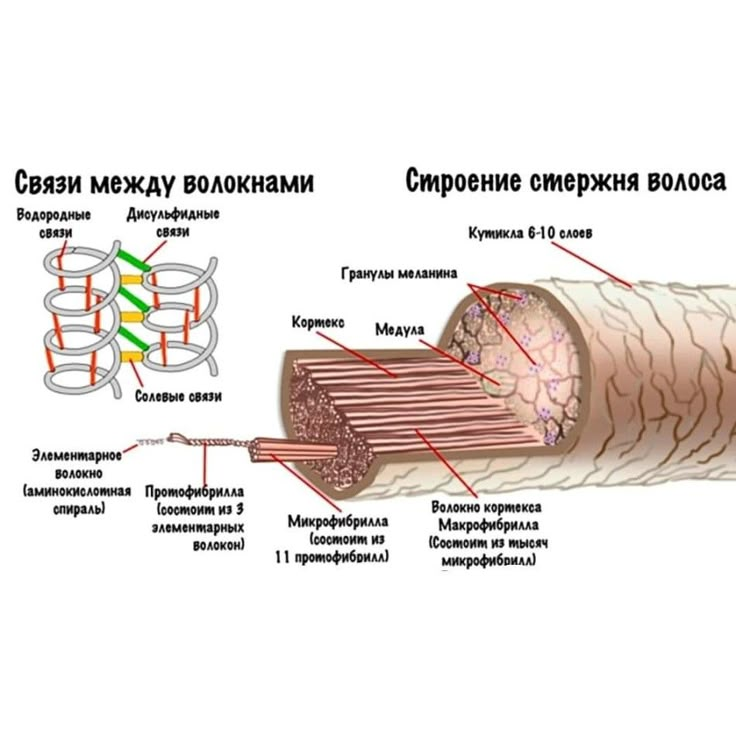 Достижения в области химии белков. 2005; 70:37–38. [PubMed] [Google Scholar]
Достижения в области химии белков. 2005; 70:37–38. [PubMed] [Google Scholar]
Mercer (1953) Mercer EH. Неоднородность кератиновых волокон. Журнал текстильных исследований. 1953; 23: 388–397. doi: 10.1177/004051755302300603. [CrossRef] [Google Scholar]
Mills et al. (2009 г.) Миллс Т.Т., Хуанг Дж., Фейгенсон Г.В., Нэгл Дж.Ф. Влияние холестерина и ненасыщенного липида доппероксидазы на упаковку цепей насыщенных бислоев дппк в гелевой фазе. Общая физиология и биофизика. 2009; 28: 126–139. doi: 10.4149/gpb_2009_02_126. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Mills et al. (2008) Миллс Т.Т., Тумбес Г.Е.С., Тристрам-Нэгл С., Смилгис Д.М., Фейгенсон Г.В., Нэгл Дж.Ф. Параметры порядка и области в жидкофазно-ориентированных липидных мембранах с использованием широкоугольного рассеяния рентгеновских лучей. Биофизический журнал. 2008;95: 669–681. doi: 10.1529/biophysj.107.127845. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Nussbaum et al. (2007) Nussbaum RL, McInnes RR, Willard HF, Hamosh A. Принципы молекулярного заболевания: уроки гемоглобинопатий. Генетика Томпсона и Томпсона в медицине. 2007;т. 6: 181–202. [Google Scholar]
(2007) Nussbaum RL, McInnes RR, Willard HF, Hamosh A. Принципы молекулярного заболевания: уроки гемоглобинопатий. Генетика Томпсона и Томпсона в медицине. 2007;т. 6: 181–202. [Google Scholar]
Охта и др. (2005) Охта Н., Ока Т., Иноуэ К., Яги Н., Като С., Хатта И. Структурный анализ комплекса клеточной мембраны волосяного волокна с помощью микролучевой рентгеновской дифракции. Журнал прикладной кристаллографии. 2005; 38: 274–279.. doi: 10.1107/S002188980403403X. [CrossRef] [Google Scholar]
Pan et al. (2008) Пан Дж., Миллс Т.Т., Тристрам-Нэгл С., Нэгл Дж.Ф. Холестерин неуниверсально возмущает липидные бислои. Письма о физическом обзоре. 2008;100:198103. doi: 10.1103/PhysRevLett.100.198103. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Pauling & Corey (1950) Pauling L, Corey RB. Две спиральные конфигурации полипептидной цепи, связанные водородными связями. Журнал Американского химического общества. 1950;72:5349–5349. doi: 10.1021/ja01167a545. [CrossRef] [Google Scholar]
Pauling & Corey (1951) Pauling L, Corey RB. Структура волос, мышц и связанных с ними белков. Труды Национальной академии наук Соединенных Штатов Америки. 1951; 37: 261–271. doi: 10.1073/pnas.37.5.261. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Структура волос, мышц и связанных с ними белков. Труды Национальной академии наук Соединенных Штатов Америки. 1951; 37: 261–271. doi: 10.1073/pnas.37.5.261. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Petrache et al. (1998) Петраче Х.И., Гуляев Н., Тристрам-Нагл С., Чжан Р., Сутер Р.М., Нэгл Дж.Ф. Межбислойные взаимодействия в рентгеновском рассеянии высокого разрешения. Физический обзор E. 1998;57:7014–7024. doi: 10.1103/PhysRevE.57.7014. [CrossRef] [Google Scholar]
Пинто и др. (2014) Пинто Н., Ян Ф.К., Негиши А., Райнштедтер М.С., Гиллис Т.Э., Фадж Д.С. Самосборка повышает прочность волокон, состоящих из белков промежуточных филаментов виментина. Биомакромолекулы. 2014; 15: 574–581. doi: 10.1021/bm401600a. [PubMed] [CrossRef] [Google Scholar]
Poinapen et al. (2013) Poinapen D, Toppozini L, Dies H, Brown DCW, Rheinstädter MC. Статические магнитные поля улучшают порядок липидов в нативной плазматической мембране растений. Мягкая материя. 2013;9: 6804–6813. doi: 10.1039/c3sm50355k. [CrossRef] [Google Scholar]
doi: 10.1039/c3sm50355k. [CrossRef] [Google Scholar]
Rafik, Doucet & Briki (2004) Rafik MEr, Doucet J, Briki F. Архитектура промежуточных филаментов, определенная с помощью рентгеновского дифракционного моделирования твердого α -кератина. Биофизический журнал. 2004; 86: 3893–3904. doi: 10.1529/biophysj.103.034694. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Randebrook (1964) Randebrook RJ. Neue erkenntnisse über den morphologischen aufbau des menschlichen haares. Журнал Общества косметических химиков. 1964;15:691–706. [Google Scholar]
Rappolt & Rapp (1996) Rappolt M, Rapp G. Одновременная мало- и широкоугольная дифракция рентгеновских лучей во время основного перехода димиристоилфосфатидилэтаноламина. Berichte der Bunsengesellschaft и Physikalische Chemie. 1996; 7: 1153–1162. doi: 10.1002/bbpc.19961000710. [CrossRef] [Google Scholar]
Rheinstädter et al. (2008) Rheinstädter MC, Das J, Flenner EJ, Brüning B, Seydel T, Kosztin I.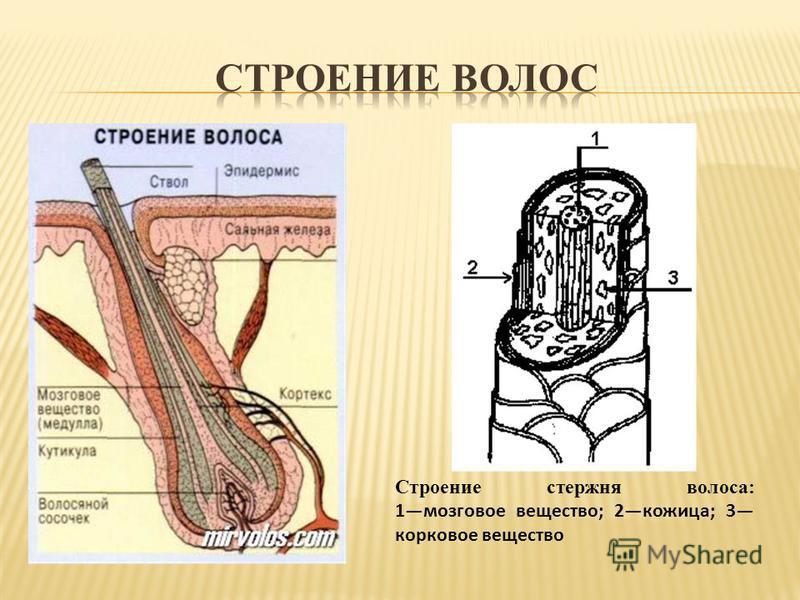 Когерентность движения в жидких фосфолипидных мембранах. Письма о физическом обзоре. 2008;101:248106. doi: 10.1103/PhysRevLett.101.248106. [PubMed] [CrossRef] [Академия Google]
Когерентность движения в жидких фосфолипидных мембранах. Письма о физическом обзоре. 2008;101:248106. doi: 10.1103/PhysRevLett.101.248106. [PubMed] [CrossRef] [Академия Google]
Rheinstädter et al. (2004) Rheinstädter MC, Ollinger C, Fragneto G, Demmel F, Salditt T. Коллективная динамика липидных мембран, изученная методом неупругого рассеяния нейтронов. Письма о физическом обзоре. 2004;93:108107. doi: 10.1103/PhysRevLett.93.108107. [PubMed] [CrossRef] [Google Scholar]
Rheinstädter, Seydel & Salditt (2007) Rheinstädter MC, Seydel T, Salditt T. Наносекундная молекулярная релаксация в липидных бислоях, изученная с помощью рассеяния нейтронов с высоким энергетическим разрешением и дифракции на месте. Физическое обозрение Е. 2007; 75:011907. doi: 10.1103/PhysRevE.75.011907. [PubMed] [CrossRef] [Google Scholar]
Robbins (2012) Robbins CR. Химическое и физическое поведение человеческих волос. 5-е изд. Нью-Йорк: Спрингер; 2012. [Google Scholar]
Rogers (1959) Rogers GE. Электронная микроскопия шерсти. Журнал ультраструктурных исследований. 1959; 2: 309–330. doi: 10.1016/S0022-5320(59)80004-6. [PubMed] [CrossRef] [Google Scholar]
Электронная микроскопия шерсти. Журнал ультраструктурных исследований. 1959; 2: 309–330. doi: 10.1016/S0022-5320(59)80004-6. [PubMed] [CrossRef] [Google Scholar]
Schneggenburger et al. (2011) Schneggenburger P, Beerlink A, Weinhausen B, Salditt T, Diederichsen U. Спирали пептидной модели в липидных мембранах: вставка, расположение и ответ липидов на агрегацию, изученные с помощью рентгеновского рассеяния. Европейский биофизический журнал. 2011;40:417–436. дои: 10.1007/s00249-010-0645-4. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Swift & Smith (2001) Swift JA, Smith JR. Микроскопические исследования эпикутикулы кератиновых волокон млекопитающих. Журнал микроскопии. 2001; 204: 203–211. doi: 10.1046/j.1365-2818.2001.00957.x. [PubMed] [CrossRef] [Google Scholar]
Tristram-Nagle et al. (2002) Тристрам-Нэгл С., Лю Ю., Леглейтер Дж., Нэгл Дж. Ф. Структура гелевой фазы dmpc определена методом рентгеноструктурного анализа. Биофизический журнал. 2002; 83: 3324–3335. дои: 10.1016/S0006-3495(02)75333-2. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
дои: 10.1016/S0006-3495(02)75333-2. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Ward & Lundgren (1954) Ward WH, Lundgren HP. Образование, состав и свойства кератинов. Достижения в области химии белков. 1954; 9: 243–297. [PubMed] [Google Scholar]
Welti et al. (1981) Велти Р., Ринтоул Д.А., Гудсаид-Залдуондо Ф., Фелдер С., Силберт Д.Ф. Фосфолипиды гель-фазы в плазматической мембране истощенных по стеролам клеток lm мыши. Журнал биологической химии. 1981; 256:7528–7535. [PubMed] [Академия Google]
Wilk, James & Amemiya (1995) Wilk KE, James VJ, Amemiya Y. Структура промежуточных нитей человеческого волоса. Biochimica et Biophysica Acta (BBA) — общие предметы. 1995;1245:392–396. doi: 10.1016/0304-4165(95)00111-5. [PubMed] [CrossRef] [Google Scholar]
Yang et al. (2014) Ян Ф.К., Петерс Р.Д., Дайс Х., Райнштадтер М.С. Иерархическая, самоподобная структура в родном загоне для кальмаров. Мягкая материя. 2014;10:5541–5549. doi: 10.1039/C4SM00301B.